一個新磷酸酶張力蛋白樣同源物亞型的研究
2016-05-14 09:03:29張謝譚麟朱杰尹博偉李宏
中國現代醫生 2016年9期
關鍵詞:細胞增殖
張謝 譚麟 朱杰 尹博偉 李宏
[摘要] 目的 鑒定新發現磷酸酶張力蛋白樣同源物(phosphatase and tensin homolog,PTEN)亞型的生物活性及潛在的腫瘤抑制作用。 方法 蛋白印跡法、免疫共沉淀法檢測各細胞株PTEN、PTEN亞型(upper PTEN,UPP)的表達;細胞轉染法轉染PTEN、UPP、空載體于HepG2和PC3細胞,細胞增殖法檢測PTEN、UPP對各株癌細胞生長情況的影響。 結果 UPP是一種緊密依賴于PTEN而存在的蛋白。UPP不是PTEN蛋白翻譯后修飾產物,可能是PTEN蛋白選擇性剪接體。與轉染空載體相比,轉染UPP和PTEN于HepG2和PC3細胞,細胞增殖明顯抑制,差異有統計學意義(P<0.05)。 結論 UPP可能是PTEN蛋白選擇性剪接產物,可能是一個新的腫瘤抑制因子。
[關鍵詞] 磷酸酶張力蛋白樣同源物;腫瘤抑制因子;細胞增殖;蛋白印跡法;選擇性剪接體
[中圖分類號] R73 [文獻標識碼] A [文章編號] 1673-9701(2016)09-0001-04
[Abstract] Objective To identify a new phosphatase and tensin homolog(PTEN) subtype and explore its potential role in tumor suppression. Methods Western blotting, immunoprecipitation were used to detect the expression of PTEN subtype(upper PTEN,UPP) and PTEN in different cell lines. PTEN, UPP and mock-vehicle were transfected to HepG2 and PC3 cell lines, then tested PTEN and UPP's effection on tumor suppression by cell proliferation bioassay. Results The expression of UPP closely depended on the existence of PTEN. UPP was not a post-translational modification product by PTEN, rather than its alternative splicing variant. Compared to cells transfected with empty vector alone, HepG2 and PC3 cells transfected with UPP or PTEN inhibited cell proliferation, and there was statistical significance(P<0.05). Conclusion UPP might be a alternative splicing variant of PTEN and a new tumor suppressor.
[Key words] Phosphatase and tensin homolog; Tumor suppressor; Cell proliferation; Western blot;Alternative splicing variant
磷酸酶張力蛋白樣同源物(phosphatase and tensin homolog,PTEN)又被稱為多種晚期癌突變基因(mutated in multiple advanced cancers,MMAC1),是繼p53之后又一個重要的腫瘤抑制因子[1-4]。我們通常所指的PTEN是一個含403個氨基酸的蛋白質,具有蛋白磷酸酶和脂質磷酸酶活性[5,6]。自從1997年PTEN被作為腫瘤抑制因子提出以來,人們發現在多種人類腫瘤中存在PTEN的缺失或突變[1-4,7]。目前針對PTEN的癌癥治療和其他疾病的治療抱有很高期望。
我們和其他實驗室發現在各種不同的組織和細胞中存在一個比PTEN大約大15kDa的同源蛋白[8,9]。此蛋白的存在嚴格依賴于PTEN的表達,且表達豐度和PTEN成正比,我們命其名為UPP(upper PTEN)。研究PTEN基因5'-UTR全序列發現在第513位核糖核苷酸存在一個非常規翻譯起始位點CTG,且UPP在N端比PTEN多173氨基酸[8-10]。本實驗擬鑒定UPP與PTEN關系,以及UPP潛在腫瘤抑制作用。
1 材料與方法
1.1 主要試劑及細胞株
DMEM高糖細胞培養液、胎牛血清(FBS)、0.25%胰蛋白酶、雙抗(100 U/mL青霉素,100U/mL鏈霉素)均購自美國Gibico公司;Anti-Human PTEN(clone 6H2.1)抗體購自美國CASCADE生物公司;α-Actin羊單克隆抗體、GAPDH小鼠單克隆抗體、兔抗人IgG、猴抗羊IgG及羊抗鼠IgG二抗均購自美國Santa Cruz公司;Lipofectamine 2000 轉染試劑盒購自美國Invitrogen公司。A431、Cos-1、LNCaP、NIH3T3、PC3、293T、U2OS、HCT116、MDA-468、HepG2細胞株均購自上海中科院細胞庫。MEF(PTEN+/+)、MEF(PTEN-/-)細胞株以及pcDNA3.1-PTEN、pcDNA3.1-UPP、空載體質粒由美國紐約斯隆癌癥中心贈送。
1.2 細胞培養及轉染
A431、Cos-1、LNCaP、PC3、293T、U2OS、HCT116、NIH3T3、MDA-468、HepG2、MEF(PTEN+/+)、MEF(PTEN-/-)細胞株用含10%FBS,雙抗(100 U/mL青霉素,100 U/mL鏈霉素)的高糖 DM EM培養基,在37℃、CO2體積分數為5%及飽和濕度條件下培養,取對數生長期細胞進行實驗。2×105細胞/3 mL接種于100 mm培養皿中,等其貼壁長到全皿的80%~90%時收蛋白。
按照Lipofectamine 2000轉染試劑盒說明書將UPP、PTEN質粒和空載體分別轉染進入HepG2細胞。轉染前1 d,細胞鋪板于100 mm培養皿中,使其轉染日密度為90%。轉染試劑用Opti-MEM無血清培養基稀釋,細胞也用Opti-MEM無血清培養基孵育。轉染6 h后,轉染培養基換成完全培養基包含10%FBS和抗生素。細胞轉染48 h后,提取蛋白。
1.3 蛋白印跡法(Western blot)檢測
收集對數生長期的各細胞株,分別用含蛋白酶抑制劑的RIPA細胞裂解液裂解,提取蛋白并定量。取50 μg總蛋白進行SDS-PAGE凝膠電泳,轉膜后以5%脫脂牛奶封閉30 min,在1∶1000濃度的Anti-Human PTEN (clone 6H2.1) 抗體及1∶400濃度 α-Actin、GAPDH抗體中室溫孵育2 h,TBST洗滌3次以后,1∶2000濃度的兔抗人IgG以及1∶1000猴抗羊IgG、羊抗鼠IgG二抗室溫孵育1.5 h,TBST洗滌3次,通過加入BeyoECL Plus顯色液于凝膠成像儀器曝光。
1.4 免疫共沉淀(Immunoprecipitation)檢測
收集對數生長期的細胞株,同上述細胞裂解后,加入3 μg PTEN抗體,4℃孵育過夜。將預處理的20 μL protein A 瓊脂糖珠加入到和抗體孵育過夜的細胞裂解液中,4℃孵育2 h。免疫沉淀反應后,瓊脂糖珠用裂解液清洗3次,PBS清洗1次,SDS-PAGE分離,western blot分析。
1.5 細胞增殖檢測
UPP、PTEN和空載體轉染 HepG2、PC3細胞,G418篩選數周后獲得穩定表達細胞。將2×105個穩定表達細胞種在細胞培養皿中,每種表達細胞各種15個培養皿。從第2天開始每天取3個培養皿用胰蛋白酶消化收集細胞、計數取平均值,連續5 d。所得平均值以天為橫坐標制作生長曲線,通過比較各種相應質粒轉染的細胞生長曲線判斷UPP、PTEN對細胞生長的影響。
1.6 統計學方法
采用SPSS13.0統計學軟件進行分析,計量資料以均數±標準差(x±s)表示,組間比較采用單因素方差分析,P<0.05為差異有統計學意義。
2 結果
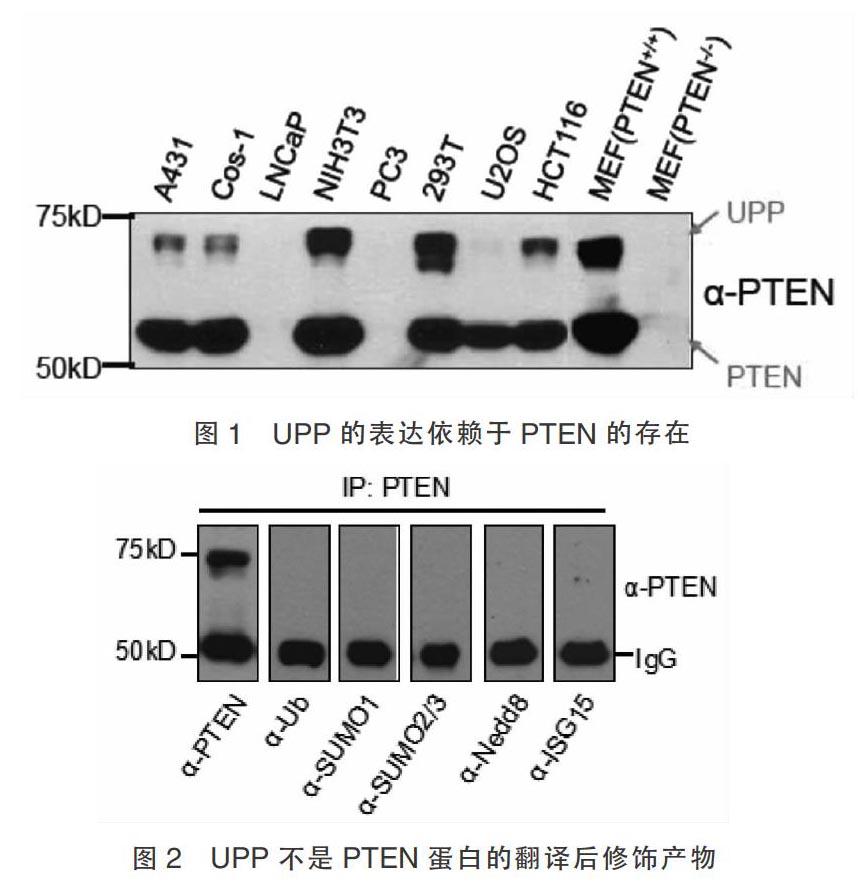
2.1 UPP是一種緊密依賴于PTEN而存在的蛋白
利用PTEN特異性抗體并借助免疫共沉淀和蛋白免疫印跡法,結果顯示在A431、Cos-1、NIH3T3、293T、U2OS、HCT116、MEF(PTEN+/+)細胞中存在一個比PTEN約大15kDa的同源蛋白,此蛋白的存在嚴格依賴于PTEN的表達,且表達豐度和PTEN成正比。其中U2OS細胞存在UPP蛋白,但是表達量低。而LNCaP、PC3、MEF(PTEN-/-)細胞中UPP和PTEN均不表達(圖1)。
2.2 UPP不是PTEN蛋白翻譯后修飾產物
根據UPP分子量的大小,我們猜測了幾種可能的翻譯后修飾,如單泛素化、SUMO修飾、Nedd8修飾、ISG15修飾,并加以驗證。在293T細胞中應用免疫沉淀法結果顯示UPP不表達,提示UPP不是PTEN基因翻譯后修飾產物(圖2)。
2.3 UPP可能是PTEN蛋白選擇性剪接產物
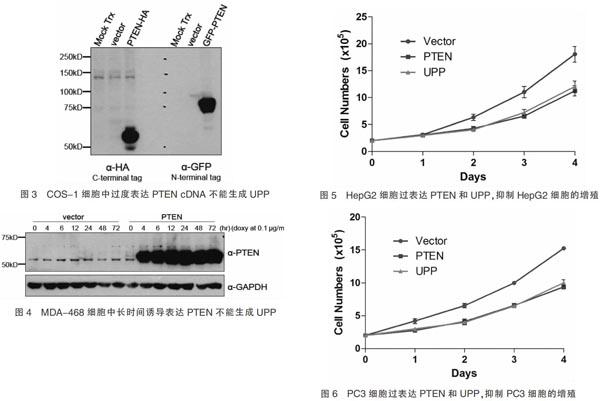
為了進一步確認UPP與PTEN的關系,我們轉染C-末端HA-,和N-末端 GFP-的PTEN cDNA于COS-1細胞,使COS-1細胞過度表達PTEN,western blot結果顯示PTEN表達量增多,但是不能檢測到UPP(圖3)。
0.1 μg/mL多西環素誘導PTEN缺失的MDA-468細胞,使其PTEN過表達,western blot結果顯示,PTEN隨著誘導時間表達量增多,但是仍無法檢測到UPP表達(圖4),以上結果說明UPP很可能是PTEN基因選擇性剪接產物。
2.4 UPP可抑制癌細胞的生長
UPP、PTEN和空載體轉染 HepG2細胞,檢測細胞增殖數。結果顯示,轉染后第1天,UPP、PTEN、空載體組細胞增殖數分別為(2.97±0.36)×105、(3.03±0.31)×105、(2.97±0.12)×105,各組間差異無統計學意義(F=0.17,P=0.85)。轉染后第2天,UPP、PTEN、空載體組細胞增殖數分別為(4.03±0.57)×105、(4.27±0.68)×105、(6.33±1.03)×105,各組間差異有統計學意義(F=7.84,P=0.02);轉染后第3天,UPP、PTEN、空載體組細胞增殖數分別為(7.23±0.96)×105、(6.57±0.68)×105、(11.03±1.80)×105,各組間差異有統計學意義(F=11.29,P=0.01);轉染后第4天,UPP、PTEN、空載體組細胞增殖數分別為(12.07±1.77)×105、(11.27±1.68)×105、(18.07±2.52)×105,各組間差異有統計學意義(F=10.13,P=0.01)(圖5)。
UPP、PTEN和空載體轉染PC3細胞,檢測細胞增殖數,結果顯示,轉染后第1天,UPP、PTEN、空載體組細胞增殖數分別為(2.99±0.30)×105、(2.75±0.42)×105、(4.18±0.67)×105,各組間差異有統計學意義(F=7.52,P=0.02)。轉染后第2天,UPP、PTEN、空載體組細胞增殖數分別為(3.97±0.60)×105、(4.11±0.66)×105、(6.53±0.55)×105,各組間差異有統計學意義(F=17.06,P<0.01);轉染后第3天,UPP、PTEN、空載體組細胞增殖數分別為(6.48±0.40)×105、(6.56±0.53)×105、(9.97±0.50)×105,各組間差異有高度統計學意義(F=52.23,P<0.01);轉染后第4天,UPP、PTEN、空載體組細胞增殖數分別為(10.01±0.78)×105、(9.38±0.58)×105、(15.26±0.48)×105,各組間差異有統計學意義(F=79.92,P<0.01)(圖6)。
3 討論
PTEN作為繼p53之后又一個重要的腫瘤抑制因子,學者針對其癌癥治療和其他疾病的治療抱有很高期望。在針對PTEN的抗癌藥物開發過程中,PTEN的調控是國內外研究的一個熱點。已知的PTEN調控方式包括氧化、乙酰化、磷酸化、泛素化等[11-14]。其中磷酸化被研究得最多。普遍存在的蛋白激酶CK2能磷酸化PTEN蛋白的C端Ser/Thr殘基[15]。PTEN去磷酸化后,其羥基端C2結構域外露,有助于PTEN實現膜轉移,進而發揮其脂質磷酸酶的功能,抑制PI3K-AKT信號通路[16-18]。去磷酸化后的PTEN相對不穩定,在執行完功能后通常通過泛素-蛋白酶體途徑被降解掉,但是CK2磷酸化的對象廣泛,雖然通過抑制CK2能夠激活PTEN,但因為其他CK2底物也受影響,副作用大[19]。
PTEN另一個被重點研究的翻譯后修飾是泛素化,泛素化后的PTEN經由蛋白酶體降解[20,21]。臨床顯示很多癌癥樣本中的PTEN泛素化酶水平偏高,說明泛素化介導的PTEN降解可能是癌癥發生的一個重要原因[20-22]。有趣的是,PTEN泛素化不僅導致它的降解。除了常見的多泛素化修飾PTEN還可被單泛素化修飾。Pier Paolo Pandolfi 實驗室報道PTEN的單泛素化幫助PTEN實現細胞核轉運[22]。臨床上,從Cowden syndrome患者中發現的PTEN突變體K13E和K289E(兩個重要的泛素化位點)雖然保留完整的脂質磷酸酶活性,卻因為不能被單泛素化,無法轉運入核行使抑癌功能[23]。這種單泛素化和多泛素化的矛盾給以PTEN泛素化酶為靶點開發癌癥新藥物帶來困惑。
利用多種PTEN特異性抗體并借助蛋白免疫印跡法,我們研究顯示在各種不同的組織和細胞中存在一個比PTEN大約大15kDa的同源蛋白——UPP。根據UPP分子量的大小,我們猜測了幾種可能的翻譯后修飾,如單泛素化、SUMO修飾、Nedd8修飾、ISG15修飾,并加以驗證。這幾種可能都被一一排除。用PTEN的cDNA轉染細胞瞬間表達或在PTEN缺失細胞中長時間誘導PTEN的表達不能產生UPP,說明UPP很可能是PTEN基因轉錄選擇性剪接產生的同源蛋白。
Hopkins等[9]是最早報道關于PTEN的亞基,他們將其命名為PTEN-Long,其具有膜脂質磷酸酶滲透性,由細胞分泌,并能進入其他細胞[9]。PTEN-Long的變異翻譯起始于上游519bp位點的CUG,N端比PTEN多173個氨基酸,與我們實驗室發現的UPP一樣。研究顯示PTEN-Long與PTEN一樣可以通過調控PI3K-AKT途徑誘導腫瘤細胞凋亡。之后Liang等[24]又發現一個PTEN亞基,命名為PTENα,其位于線粒體,與PTEN一起調控線粒體能量代謝。PTENα也是另一種翻譯剪接體,但是其翻譯也起源于上游一個非常規翻譯密碼CUG,其N段比PTEN多了一個173氨基酸(人類)或者169個氨基酸(小家鼠類)。但是PTEN-Long和PTENα調控的是兩個不同的生物學途徑,故他們可能是兩個不同的PTEN亞基。
轉染UPP于HepG2肝癌細胞可抑制其增殖,說明UPP抑制癌細胞生長。通常認為通過腺病毒將PTEN的cDNA重新引入有PTEN基因缺陷的患者體內,可以抑制因PI3K-AKT通路過于活躍導致的癌癥[25]。但是,越來越多的臨床報道顯示腺病毒介導的基因治療存在安全隱患,接受基因治療的患者并沒有像所期望的那樣從癌癥中康復,有相當一部分患者出現了由腺病毒導致的各類副作用包括新的癌癥。新發現的UPP蛋白可能可以規避以PTEN基因治療帶來的副作用,為針對UPP設計治療癌癥和其他PTEN相關疾病的新方法提供依據。
[參考文獻]
[1] Li J,Yen C,Liaw D,et al. PTEN,a putative protein tyrosine phosphatase gene mutated in human brain,breast,and prostate cancer[J]. Science,1997,275(5308):1943-1947.
[2] Milelam I,Falcone I,Conciatori F,et al. PTEN:Multiple functions in human malignant tumors[J]. Front Oncol,2015, 5:24.
[3] Garcia-Cao I,Song MS,Hobbs RM,et al. Systemic elevation of PTEN induces tumor-suppressive metabolic state[J].Cell,2012,149(1):49-62.
[4] Liu C,Li G,Chen R,et al. A novel PTEN gene promoter mutation and untypical Cowden syndrome[J]. Chin J Cancer Res,2013,25(3):306-311.
[5] Maehama T,Dixon JE. The tumor sPTEN-Longressor,PTEN/MMAC1,dephosphorylates the lipid second messenger,phosphatidylinositol 3,4,5-trisphosphate[J]. J Biol Chem,1998,273(22):13375-133758.
[6] Leslie NR,Macario H,Spinelli L,et al. The significance of PTEN's protein phosphatase activity[J]. Adv Enzyme Regul,2009,49(1):190-196.
[7] Ahearn TU,Pettersson A,Ebot EM,et al. A prospective investigation of PTEN loss and ERG expression in lethal prostate cancer[J]. J Natl Cancer Inst,2015,108(2).
[8] Wang H,Zhang P,Lin C,et al. Relevance and therapeutic possibility of PTEN-long in renal cell carcinoma[J]. PLoS One,2015,10(2):e114250.
[9] Hopkins BD,Fine B,Steinbach N,et al. A secreted PTEN phosphatase that enters cells to alter signaling and survival[J]. Science,2013,341(6144):399-402.
[10] Masson GR,Perisic O,Burke JE,et al. The intrinsically disordered tails of PTEN and PTEN-L have distinct roles in regulating substrate specificity and membrane activity[J].Biochem J,2016,473(2):135-144.
[11] Chetram MA,Don-Salu-Hewage AS,Hinton CV. ROS enhances CXCR4-mediated functions through inactivation of PTEN in prostate cancer cells[J]. Biochem Bioph Res Co,2011,410(2):195-200.
[12] Ikenoue T,Inoki K,Zhao B,et al. PTEN acetylation modulates its interaction with PDZ domain[J]. Cancer Res,2008,68(17):6908-6912.
[13] Dida F,Li Y,Iwao A,et al. Resistance to TRAIL-induced apoptosis caused by constitutional phosphorylation of Akt and PTEN in acute lymphoblastic leukemia cells[J]. Exp Hematol,2008,36(10):1343-1353.
[14] Maddika S. WWP2 is an E3 ubiquitin ligase for PTEN[J].Nature Cell Biology,2011,13(6):728-733.
[15] Shi Y,Paluch BE,Wang X,et al. PTEN at a glance[J]. J Cell Sci,2012,125(Pt 20):4687-4692.
[16] Vazquez F,Grossman SR,Takahashi Y,et al. Phosphorylation of the PTEN tail acts as an inhibitory switch by preventing its recruitment into a protein complex[J]. J Biol Chem,2001, 276(52):48627-48630.
[17] Pérez-Ramírez C,Ca?觡adas-Garre M,Molina MA,et al. PTEN and PI3K/AKT in non-small-cell lung cancer[J]. Pharmacogenomics,2015,6(16):1843-62.
[18] Carnero A,Paramio JM. The PTEN/PI3K/AKT pathway in vivo,cancer mouse models[J]. Front Oncol,2014,4:252.
[19] Torres,Pulido R. The tumor suppressor PTEN is phosphor- ylated by the protein kinase CK2 at its C terminus. Implications for PTEN stability to proteasome-mediated degradation[J]. J Biol Chem,2001,276(2):993-998.
[20] Wang X,Jiang X. PTEN:A default gate-keeping tumor suppressor with a versatile tail[J]. Cell Res,2008,18(8):807-816.
[21] Errafiy R,Aguado C,Ghislat G,et al. PTEN increases autophagy and inhibits the ubiquitin-proteasome pathway in glioma cells independently of its lipid phosphatase activity[J]. PLoS One,2013,8(12):e83318.
[22] Song MS,Salmena L,Pandolfi PP. The functions and regulation of the PTEN tumour suppressor[J]. Nat Rev Mol Cell Bio,2012,13(5):283-296.
[23] Trotman LC,Wang X,Alimonti A,et al. Ubiquitination regulates PTEN nuclear import and tumor suppression[J]. Cell,2007,128(1):141-156.
[24] Liang H,He S,Yang J,et al. PTENα,a PTEN isoform translated through alternative initiation,regulates mitochondrial function and energy metabolism[J]. Cell Metab,2014,19(5):836-848.
[25] Liang H,He S,Yang J,et al. PTEN activation sensitizes breast cancer to PI3-kinase inhibitor through the β-catenin signaling pathway[J]. Oncol Rep,2012,28(3):943-948.
(收稿日期:2015-12-11)
猜你喜歡
中國當代醫藥(2017年2期)2017-03-18 18:17:33
右江醫學(2016年4期)2017-01-05 16:26:48
中國民族民間醫藥·下半月(2016年6期)2016-11-02 20:27:22
中國醫藥科學(2016年9期)2016-07-25 23:04:03
湖南師范大學學報·自然科學版(2016年3期)2016-06-25 06:18:59
學周刊·下旬刊(2016年7期)2016-05-20 18:48:49
課程教育研究·學法教法研究(2016年6期)2016-04-26 10:06:57
新課程·中學(2015年9期)2015-10-26 19:00:20
