甜椒核心種質遺傳多樣性與親緣關系分析
2016-07-23 14:32:36劉子記申龍斌楊衍
江蘇農業科學 2016年5期
關鍵詞:相關性
劉子記++申龍斌++楊衍

摘要:以102份甜椒核心種質為研究材料,對8個農藝性狀進行遺傳多樣性分析,并且基于基因型效應值進行性狀間的相關性和種質間的親緣關系分析。8個農藝性狀的多樣性指數分別為4.60、4.60、4.59、4.57、4.60、4.58、4.58和4.53,表明甜椒核心種質存在豐富的遺傳多樣性。首花節位與株高呈極顯著正相關,相關系數為0.38;果縱徑與株高呈極顯著正相關,與首花節位呈顯著相關,相關系數分別為0.37和0.23;果柄長與株高呈顯著相關,相關系數為020;果肉厚與果縱徑、果橫徑呈極顯著正相關,相關系數分別為0.28和0.30;單果質量與果縱徑、果橫徑、果肉厚呈極顯著正相關,與首花節位呈顯著正相關,相關系數分別為0.37、0.67、0.53和0.20。不同種質間遺傳距離變幅為151~10.41,表明這些核心材料間遺傳差異較大。在聚類重新標定距離為16.5時,102份甜椒核心種質被分為27個類群,其中20個類群分別由1份種質材料組成,表明這20份種質與其余種質親緣關系較遠。該研究明確了不同甜椒種質間的親緣關系,為甜椒核心種質資源的有效利用和新品種選育提供了理論依據。
關鍵詞:甜椒;基因型值;遺傳多樣性;相關性;親緣關系
中圖分類號: S641.303文獻標志碼: A文章編號:1002-1302(2016)05-0199-03
收稿日期:2015-04-22
基金項目:中國熱帶農業科學院院本級基本科研業務費專項(編號:1630032015003)。
作者簡介:劉子記(1982—),男,博士,副研究員,研究方向:蔬菜分子生物學及遺傳育種。E-mail:liuziji1982@163.com。
通信作者:曹振木,研究員,研究方向為蔬菜遺傳育種。E-mail:catasvegetable@126.com。甜椒(Capsicum annuum var. grossum,2n=2x=24)原產于中南美洲,在植物學分類上屬于茄科(Solanaceae)辣椒屬(Capsicum)[1]。辣椒屬包括5個栽培種[2],甜椒屬于一年生辣椒栽培種。甜椒果實不辣而略帶甜味,大量研究表明甜椒果實中含有豐富的維生素A[3]、維生素C[4]、類胡蘿卜素[5]及礦物質,可以保護生物有機體免受氧化傷害[6-7]、提高機體免疫力和預防壞血病。甜椒以果實特有的色澤和營養成分成為一種世界性的蔬菜作物。作物種質資源為栽培種遺傳改良、新品種選育及遺傳學研究提供豐富的遺傳變異和基因資源[8]。加速新品種的選育和推廣利用是科技興農的關鍵,種質資源是實現各個育種途徑的原始材料,對于原始材料的選擇依賴于所掌握種質資源的廣度及對其研究的深度[9]。親緣關系分析是植物種質資源研究的主要內容之一。開展作物種質資源親緣關系研究,有助于了解材料的遺傳背景,為種質資源的創新利用與新品種選育提供重要信息[10]。以往辣椒種質資源親緣關系分析多集中在有辣味的辣椒材料[11-15],有關甜椒核心種質親緣關系的研究鮮有報道。
作物種質的遺傳多樣性是其適應環境變化的表現,表型性狀的遺傳多樣性研究為從整體上評價和利用種質資源奠定基礎。但由于種質資源的形態特征是基因型、環境以及基因型與環境互作的綜合表現[16],僅根據農藝性狀表型值難以鑒定其遺傳背景的異同。為了排除環境條件、基因型與環境互作的影響,準確度量不同遺傳材料間的遺傳差異,本研究采用混合線性模型無偏預測性狀的基因型值,基于基因型值進行性狀間的相關性分析和材料間的親緣關系分析,以期為甜椒種質資源的收集及遺傳育種提供參考。
1材料與方法
1.1試驗材料和基因型值預測
將102份甜椒核心種質按隨機區組設計種植于中國熱帶農業科學院熱帶作物品種資源研究所10隊試驗基地,3次重復,參考《辣椒種質資源描述規范與數據標準》調查株高、株幅、首花節位、果縱徑、果橫徑、果柄長、果肉厚、單果質量。采用朱軍提出的混合線性模型無偏預測性狀的基因型值[17]。
1.2遺傳多樣性和相關性分析
采用SAS 9.0軟件分析8個性狀的最小值、最大值、平均值、極差、變異系數、方差和遺傳多樣性指數H′,基于性狀的基因型值計算各性狀之間的相關性系數。
1.3聚類分析
采用SPSS 9.0軟件基于8個性狀的基因型值對102份甜椒核心種質構建聚類圖,樣本間的遺傳距離采用歐氏距離法計算,歐氏距離計算公式為EUCLID=∑ki=1(xi-yi)2,其中,k表示樣本有k個變量,xi表示第1個樣本在第i個變量上的取值,yi標示第2個樣本在第i個變量上的取值,樣本與小類之間的遺傳距離采用最短距離法進行計算[18]。
2結果與分析
2.1甜椒農藝性狀的遺傳多樣性分析
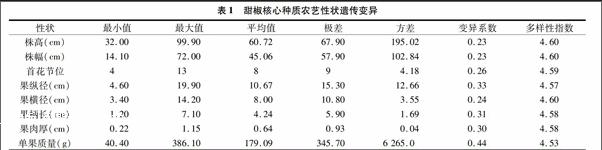
株幅、果縱徑和單果質量的極差分別為57.90 cm、15.30 cm 和345.70 g,尤其是單果質量的極差,幾乎是均值的2倍,遠遠大于其平均值,這說明甜椒核心種質的株幅、果縱徑和單果質量表型值更為分散。單果質量的變異系數最大,為0.44,果縱徑的變異系數次之,為0.33,進一步說明了甜椒核心種質的單果質量和果縱徑表型值的離散程度較高,各種質之間的遺傳差異較大。8個農藝性狀的多樣性指數分別為4.60、4.60、4.59、4.57、4.60、4.58、4.58和4.53,均超過了4.5,該結果表明甜椒核心種質存在豐富的遺傳多樣性(表1)。表1甜椒核心種質農藝性狀遺傳變異
性狀最小值最大值平均值極差方差變異系數多樣性指數株高(cm) 32.0099.9060.7267.90195.020.234.60株幅(cm) 14.1072.0045.0657.90102.840.234.60首花節位 413894.180.264.59果縱徑(cm) 4.6019.9010.6715.3012.660.334.57果橫徑(cm) 3.4014.208.0010.803.550.244.60果柄長(cm) 1.207.104.245.901.690.314.58果肉厚(cm) 0.221.150.640.930.040.304.58單果質量(g) 40.40386.10179.09345.706 265.00.444.53
2.2甜椒農藝性狀的相關性分析
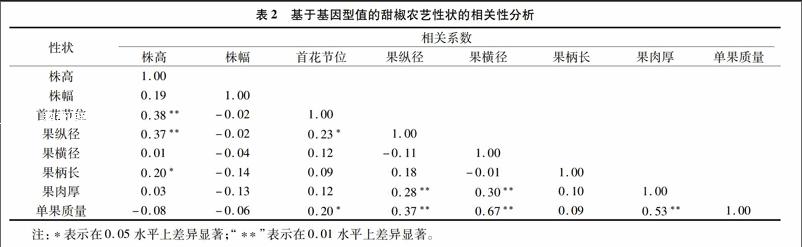
性狀的相關性可以通過對一種性狀的選擇間接達到選擇另一種性狀的效果,從而可以提高選擇效率,加速育種進程。利用復合線性模型預測102份甜椒核心種質8個農藝性狀的基因型效應值。基于基因型值進行性狀間的相關性分析,結果表明,首花節位與株高呈極顯著正相關,相關系數為0.38;果縱徑與株高呈極顯著正相關,與首花節位呈顯著相關,相關系數分別為0.37和0.23;果柄長與株高呈顯著相關,相關系數為0.20;果肉厚與果縱徑、果橫徑呈極顯著正相關,相關系數分別為0.28和0.30;單果質量與果縱徑、果橫徑、果肉厚呈極顯著正相關,與首花節位呈顯著正相關,相關系數分別為0.37、0.67、0.53和0.20(表2)。在甜椒新品種選育過程中,篩選具有較大果縱徑、果橫徑和果肉厚的育種材料可有效提高單果質量,篩選株高較低的育種材料有利于培育早熟品種。表2基于基因型值的甜椒農藝性狀的相關性分析
性狀相關系數株高株幅首花節位果縱徑果橫徑果柄長果肉厚單果質量株高1.00株幅0.191.00首花節位0.38**-0.021.00果縱徑0.37**-0.020.23*1.00果橫徑0.01-0.040.12-0.111.00果柄長0.20*-0.140.090.18-0.011.00果肉厚0.03-0.130.120.28**0.30**0.101.00單果質量-0.08-0.060.20*0.37**0.67**0.090.53**1.00注:*表示在0.05水平上差異顯著;“**”表示在0.01水平上差異顯著。
2.3甜椒核心種質聚類分析
利用歐氏距離法基于8個農藝性狀的基因型效應值計算甜椒核心材料間的遺傳距離,在供試的102份核心材料中,遺傳距離變幅為1.51~10.41,表明這些核心材料間遺傳差異較大。其中,SP165和SP106、SP106和SP76、SP225和SP177、SP214和SP206、SP196和SP214、SP165和SP219、SP106和SP257、SP219和SP160、SP131和SP177、SP206和SP68、SP165和SP76、SP106和SP79、SP76和SP257、SP219和SP106、SP225和SP131間遺傳距離較小,遺傳距離分別為1.51、158、1.60、1.62、1.69、1.76、1.78、1.79、1.81、1.83、1.84、185、1.85、1.91、1.95,表明這些材料間親緣關系相對較近。另外,SP276和SP158、SP381和SP40、SP55和SP383、SP208和SP155、SP62和SP319、SP107和SP219、SP107和SP178、SP152和SP178、SP92和SP155、SP55和SP155、SP241和SP107、SP160和SP107、SP107和SP381、SP383和SP208、SP107和SP62、SP107和SP158、SP276和SP155、SP208和SP382、SP62和SP276、SP155和SP152、SP382和SP152、SP382和SP107、SP152和SP383、SP383和SP107、SP155和SP107間遺傳距離較大,均超過了8.50,遺傳距離分別為856、8.56、8.56、8.57、8.58、8.62、8.63、8.67、8.71、8.72、878、8.79、897、9.05、9.10、9.11、9.23、9.28、9.40、9.78、983、10.01、10.03、10.28、10.41,表明這些材料間親緣關系較遠。
在聚類重新標定距離為16.5時,102份甜椒核心種質被分為27個類群,第1個類群包括2份種質,分別為SP10和SP30,表明這2份種質親緣關系相對較近;第2類群包括57份種質;第3類群包括1份種質,為SP31;第4類群包括3份種質,分別為SP36、SP50和SP40,表明這3份種質親緣關系相對較近;第5類群包括1份種質,為SP37;第6類群包括1份種質,為SP55;第7類群包括14份種質,分別為SP361、SP365、SP315、SP399、SP355、SP261、SP346、SP259、SP301、SP93、SP142、SP92、SP391和SP266;第8類群包括1份種質,為SP104;第9類群包括1份種質,為SP107;第10類群包括2份種質,分別為SP108和SP152,表明這2份種質親緣關系相對較近;第11類群包括1份種質,為SP143;第12類群包括1份種質,為SP155;第13類群包括1份種質,為SP158;第14類群包括1份種質,為SP208;第15類群包括1份種質,為SP237;第16類群包括1份種質,為SP270;第17類群包括1份種質,為SP275;第18類群包括1份種質,為SP276;第19類群包括2份種質,分別為SP279和SP368;第20類群包括2份種質,分別為SP285和SP359;第21類群包括1份種質,為SP314;第22類群包括1份種質,為SP318;第23類群包括1份種質,為SP354;第24類群包括1份種質,為SP381;第25類群包括1份種質,為SP382;第26類群包括1份種質,為SP383;第27類群包括1份種質,為SP393。從聚類結果可以看出,甜椒核心種質間遺傳差異顯著,存在豐富的遺傳多樣性,第3、5、6、8、9、11、12、13、14、15、16、17、18、21、22、23、24、25、26、27類群分別由1份種質材料組成,表明這20份種質與其余種質親緣關系較遠(圖1)。
3討論
作物種質資源的收集與保存對新品種選育、優異基因發掘以及種質創新具有重要意義[19]。世界各國相繼建立了不同作物的種質資源庫。隨著種質資源的不斷收集,種質庫變得越來越大,極大地提高了種質資源的管理費用,增加了特異種質篩選發掘的難度[20]。在前期的工作中,筆者所在研究室從410份甜椒種質中抽取102份材料構建了甜椒核心種質庫。對作物種質資源進行遺傳多樣性研究,可為育種工作提供重要的信息。本研究分析了102份甜椒核心種質8個農藝性狀的遺傳多樣性,8個農藝性狀的多樣性指數均超過了45,該結果表明甜椒核心種質存在豐富的遺傳多樣性。
作物的農藝性狀間往往存在錯綜復雜的相互關系。相關性分析能對不同數量性狀兩組變量間進行相關性研究,通過對一種性狀的選擇達到改良另一種性狀的效果,這對于不容易鑒定的數量性狀顯得尤為重要,為開展多個數量性狀綜合選擇提供依據[21]。本研究基于102份甜椒核心種質8個農藝性狀的基因型值進行性狀間的相關性分析,結果表明,首花節位與株高呈極顯著正相關;果縱徑與株高呈極顯著正相關,與首花節位呈顯著相關;果柄長與株高呈顯著相關;果肉厚與果縱徑、果橫徑呈極顯著正相關;單果質量與果縱徑、果橫徑、果肉厚呈極顯著正相關,與首花節位呈顯著正相關。
種質資源的性狀特征受基因型、環境以及基因型與環境互作的影響[22],單純依靠農藝性狀數據進行親緣關系分析存在一定誤差。如何準確地評價不同材料在遺傳上的差異則是合理分析種質親緣關系的前提。本研究采用混合線性模型無偏預測法預測性狀的基因型值,基于性狀的基因型值進行甜椒核心種質親緣關系分析,排除了環境、基因型與環境互作及農業試驗中不可控制的一些誤差影響,分析結果更具可靠性。在聚類重新標定距離為16.5時,102份甜椒核心種質被分為27個類群,其中20個類群分別由1份種質材料組成,從聚類結果可以看出,甜椒核心種質間遺傳差異顯著,存在豐富的遺傳多樣性。本研究明確了不同甜椒核心材料間的親緣關系,為甜椒種質資源的有效利用和新品種選育奠定了堅實基礎。
參考文獻:
[1]Kim S,Park M,Yeom S I,et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species[J]. Nature Genetics,2014,46(3):270-278.
[2]孟金貴,張卿哲,王碩,等. 涮辣與辣椒屬5個栽培種親緣關系的研究[J]. 園藝學報,2012,39(8):1589-1595.
[3]Mejia L A,Hudson E,Demejia E G,et al. Carotenoid content and vitamin-a activity of some common cultivars of Mexican peppers (Aapsicum annuum) as determined by HPLC[J]. Journal of Food Science,1988,53(5):1448-1451.
[4]Vanderslice J T,Higgs D J,Hayes J M,et al. Ascorbic acid and dehydroascorbic acid content of foods-as-eaten[J]. Journal of Food Composition and Analysis,1990,3(2):105-118.
[5]Ha S H,Kim J B,Park J S,et al. A comparison of the carotenoid accumulation in Capsicum varieties that show different ripening colours:deletion of the capsanthin-capsorubin synthase gene is not a prerequisite for the formation of a yellow pepper[J]. Journal of Experimental Botany,2007,58(12):3135-3144.
[6]Sun T,Xu Z,Wu C T,et al. Antioxidant activities of different colored sweet bell peppers (Capsicum annuum L.)[J]. Journal of Food Science,2007,72(2):S98-102.
[7]Marín A,Ferreres F,Tomás-Barberán F A,et al. Characterization and quantitation of antioxidant constituents of sweet pepper (Capsicum annuum L.)[J]. Journal of Agricultural and Food Chemistry,2004,52(12):3861-3869.
[8]朱巖芳,祝水金,李永平,等. ISSR分子標記技術在植物種質資源研究中的應用[J]. 種子,2010,29(2):55-59.
[9]譚亮萍,周火強,曾化偉,等. 辣椒種質資源鑒定、評價及利用研究進展[J]. 辣椒雜志,2008(2):24-28.
[10]王振東,陳超力,于佰雙,等. 大豆抗旱種質資源遺傳多樣性的SSR分析[J]. 大豆科學,2010,29(3):370-373.
[11]李永平,林琿,溫慶放. 辣椒種質資源的遺傳多樣性分析[J]. 福建農業學報,2011,26(5):747-752.
[12]李晴,張學時,張廣臣,等. 辣椒種質遺傳多樣性的RAPD分析[J]. 北方園藝,2010(22):118-122.
[13]何建文,楊文鵬,韓世玉,等. 貴州辣椒地方品種分子遺傳多樣性分析[J]. 貴州農業科學,2009,37(8):15-18.
[14]陳學軍,周坤華,宗洪霞,等. 中國灌木辣椒種質遺傳多樣性的SRAP和SSR分析[J]. 西北植物學報,2012,32(11):2201-2205.
[15]陳學軍,程志芳,陳勁楓,等. 辣椒種質遺傳多樣性的RAPD和ISSR及其表型數據分析[J]. 西北植物學報,2007,27(4):662-670.
[16]張嘉楠,昌小平,郝晨陽,等. 北方冬麥區小麥抗旱種質資源遺傳多樣性分析[J]. 植物遺傳資源學報,2010,11(3):253-259.
[17]朱軍. 作物雜種后代基因型值和雜種優勢的預測方法[J]. 生物數學學報,1993,8(1):32-44.
[18]裴鑫德. 多元統計分析及其應用[M]. 北京:北京農業大學出版社,1991.
[19]李長濤,石春海,吳建國,等. 利用基因型值構建水稻核心種質的方法研究[J]. 中國水稻科學,2004,18(3):218-222.
[20]徐海明,邱英雄,胡晉,等. 不同遺傳距離聚類和抽樣方法構建作物核心種質的比較[J]. 作物學報,2004,30(9):932-936.
[21]王瑞清,閆志順,劉英. 冬小麥品種數量性狀的典型相關分析[J]. 種子,2004,23(11):56-58.
[22]苗錦山,劉彩霞,戴振建,等. 蔥種質資源數量性狀的聚類分析、相關性和主成分分析[J]. 中國農業大學學報,2010,15(3):41-49.張斌,周廣柱,聶義豐,等. 干旱脅迫對銀杏幼苗葉片光合性狀的影響[J]. 江蘇農業科學,2016,44(5):202-205.
猜你喜歡
商情(2016年42期)2016-12-23 14:25:52
商情(2016年42期)2016-12-23 13:35:35
東方教育(2016年4期)2016-12-14 22:15:13
財經界·學術版(2016年19期)2016-11-16 16:28:33
科技視界(2016年21期)2016-10-17 17:37:34
中國實用醫藥(2016年24期)2016-10-17 04:31:12
中國實用醫藥(2016年24期)2016-10-17 03:37:40
中國實用醫藥(2016年24期)2016-10-17 03:35:06
科學與財富(2016年28期)2016-10-14 21:58:50
