產(chǎn)KPC酶肺炎克雷伯菌對(duì)頭孢他啶/阿維巴坦耐藥機(jī)制研究進(jìn)展
2023-04-29 00:44:03鄭媚匡徐李甫豪馮可瑩于洋
國(guó)外醫(yī)藥抗生素分冊(cè) 2023年3期
關(guān)鍵詞:突變
鄭媚 匡徐 李甫豪 馮可瑩 于洋
摘要:頭孢他啶/阿維巴坦(Ceftazidime/Avibactam,CZA)是頭孢他啶和一種新型β-內(nèi)酰胺酶抑制劑(阿維巴坦)的組合,已被美國(guó)食品藥品監(jiān)督管理局和國(guó)家市場(chǎng)監(jiān)督管理總局批準(zhǔn),主要用于治療復(fù)雜的腹部和尿道感染、醫(yī)院獲得性細(xì)菌性肺炎和呼吸機(jī)相關(guān)的細(xì)菌性肺炎,特別是由肺炎克雷伯菌引起的感染和炎癥。然而,CZA臨床應(yīng)用導(dǎo)致了多重耐藥肺炎克雷伯菌的出現(xiàn)。其耐藥機(jī)制主要有三種:blaKPC基因的突變、細(xì)菌細(xì)胞膜孔蛋白突變和KPC酶的表達(dá)量增加。本文就耐CZA的肺炎克雷伯菌流行情況和耐藥機(jī)制進(jìn)行綜述,旨在為臨床防治CZA的耐藥菌株提供科學(xué)依據(jù),并為新藥開發(fā)提供指導(dǎo)。
關(guān)鍵詞:產(chǎn)KPC酶肺炎克雷伯菌;頭孢他啶;阿維巴坦;耐藥機(jī)制;作用機(jī)理;突變
中圖分類號(hào):R978.1? ? ? ? 文獻(xiàn)標(biāo)志碼:A? ? ? ? ?文章編號(hào):1001-8751(2023)03-0160-06
Research Progress on the Resistance Mechanism of KPC-Producing Klebsiella pneumoniae to Ceftazidime/Avibactam
Zheng Mei1,? ?Kuang Xu1,? ?Li Fu-hao1,? ?Feng Ke-ying2,? ?Yu Yang1
(1 South China Agricultural University,Guangzhou 510642;2 Feipeng Biotechnology Co., Ltd,Dongguan 523808)
Abstract: Ceftazidime/avibactam (CZA) is a combination of ceftazidime and a novel β-lactamase inhibitor (avibactam) approved by the U.S. Food and Drug Administration and the State Administration of China for the treatment of complex abdominal and urinary tract infections, hospital-acquired bacterial pneumonia, and ventilator-associated bacterial pneumonia, particularly those caused by Klebsiella pneumoniae. However, the clinical use of CZA has led to the emergence of multidrug-resistant Klebsiella pneumoniae. There are three main mechanisms of drug resistance: mutation of blaKPC, mutation of cell membrane pore protein, and increased expression of KPC enzyme. The article reviews the prevalence and mechanism of Klebsiella pneumoniae resistant to CZA, aiming to provide scientific basis for the clinical prevention and treatment of drug-resistant strains of CZA and provide guidance for the development of new drugs.
Key words: KPC-producing Klebsiella pneumoniae;ceftazidime ;avibactam ;drug resistance; mechanism? mechanism of action;mutation
1 碳青霉烯類耐藥的肺炎克雷伯菌(CRKP)流行現(xiàn)狀
據(jù)估計(jì),在世界范圍內(nèi),因細(xì)菌感染引起的死亡人數(shù)每年可達(dá)700,000,預(yù)計(jì)到2050年,將有1000萬(wàn)人死于多重耐藥性微生物引起的相關(guān)疾病[1]。抗生素耐藥性已成為當(dāng)今最緊迫的公共衛(wèi)生挑戰(zhàn)之一,而肺炎克雷伯菌(Klebsiella pneumoniae)是一種重要的革蘭陰性機(jī)會(huì)致病菌,具有超強(qiáng)的環(huán)境適應(yīng)能力,除污水、土壤、地表水、工業(yè)廢水和植被外,肺炎克雷伯菌還可寄居在易感人類和動(dòng)物粘膜表面,并誘導(dǎo)局部組織感染,包括尿路感染、菌血癥、肺炎及肝膿腫等,或誘發(fā)全身性感染,如敗血癥等,甚至危及患者生命[2-3]。肺炎克雷伯菌被列為ESKAPE病原體,已成為我國(guó)僅次于大腸埃希菌的第二大革蘭陰性致病菌,其菌體細(xì)胞內(nèi)可攜帶多種高效的可移動(dòng)性抗菌藥物耐藥性維持和傳播的遺傳元件,是抗生素耐藥性(Antimicrobial Resistance,AMR)在人、動(dòng)物和環(huán)境中傳播的重要媒介。在抗菌藥物壓力下,由于這些質(zhì)粒、轉(zhuǎn)座子、插入序列等可移動(dòng)元件,肺炎克雷伯菌已進(jìn)化出多種抗菌藥耐藥性表型,且耐藥基因在種屬間及跨種屬間廣泛傳播,導(dǎo)致出現(xiàn)了具有“超級(jí)耐藥性”的泛耐藥菌株(Extensive drug resistance,XDR)[4]。碳青霉烯類耐藥的肺炎克雷伯菌(Carbapenems resistant K. pneumoniae, CRKP)在國(guó)內(nèi)外有越來(lái)越多的報(bào)道。在意大利,美國(guó)紐約州感染CRKP引起的死亡率分別為41.6%和48%。其中,ST258以及ST512是CRKP在美洲和南歐的主要克隆株[5-6]。值得注意的是,根據(jù)我國(guó)抗菌藥物監(jiān)測(cè)網(wǎng)絡(luò)(CHINET)報(bào)道顯示,在我國(guó)肺炎克雷伯菌對(duì)美羅培南和亞胺培南的耐藥率上升 8 倍以上[7]。近十年間, CRKP在我國(guó)廣泛傳播,流行范圍幾乎覆蓋了所有省份,且引起了較高的臨床發(fā)病率;其中基因型ST11是我國(guó)CRKP感染中最主要的MLST分型[8]。而ST11和ST258屬于克隆復(fù)合物CC258,其包含ST11、ST258和另外五個(gè)分型ST270、ST340、ST379、ST407和ST418 [9]。此外,新出現(xiàn)的ST307也被認(rèn)為是一種高危克隆型,其在醫(yī)院的高流行率應(yīng)該引起重視[10]。CRKP菌株中碳青霉烯酶編碼基因包括KPC、NDM、OXA、IMP、VIM,最主要的碳青霉烯酶基因是blaKPC-2,blaNDM-1,blaOXA-48,其中ST11型攜帶blaKPC-2的肺炎克雷伯菌的克隆傳播速度驚人[11]。CRKP感染通常具有較高的發(fā)病率和死亡率,給臨床治療帶來(lái)了嚴(yán)峻的挑戰(zhàn)[12 -13]。
2 CZA使用現(xiàn)狀以及作用機(jī)理
目前用于治療CRKP感染的抗生素選擇十分有限,主要包括多黏菌素、替加環(huán)素和氨基糖苷類藥物,因此亟需有效的治療CRKP感染的方法。頭孢他啶/阿維巴坦(CZA)由阿斯利康制藥有限公司開發(fā),于2015年2月獲得美國(guó)食品和藥物管理局批準(zhǔn)[14-15],并于2019年底在中國(guó)投入使用。它將廣譜頭孢菌素頭孢他啶與新型非β-內(nèi)酰胺酶抑制劑阿維巴坦結(jié)合,用于治療復(fù)雜的腹部和尿道感染、醫(yī)院獲得性細(xì)菌性肺炎和呼吸機(jī)相關(guān)的細(xì)菌性肺炎[16]。毫無(wú)疑問(wèn),CZA的出現(xiàn)為對(duì)抗CRKP感染提供了十分寶貴的替代方案。值得注意的是,盡管CZA對(duì)大多數(shù)碳青霉烯類耐藥肺炎克雷伯菌都是敏感的,但是在我國(guó)臨床使用CZA之前就已經(jīng)出現(xiàn)了對(duì)該藥物的耐藥菌[17]。有研究表明,KPC-2單個(gè)基因替換突變使得阿維巴坦對(duì)KPC的β-內(nèi)酰胺酶失去抑制作用[18],以及突變的KPC-3酶顯著降低了肺炎克雷伯菌對(duì)CZA的易感性[19]。一方面阿維巴坦可以保護(hù)頭孢他啶免受KPCs、OXA-48、AmpC等β-內(nèi)酰胺酶的水解,但無(wú)法抵抗金屬β-內(nèi)酰胺酶(MBLs),如NDM、VIM或IMP的水解。因此,阿維巴坦可恢復(fù)頭孢他啶對(duì)KPC陽(yáng)性肺炎克雷伯菌(KPC-Kp)的活性,在受到KPC酶親核進(jìn)攻后,阿維巴坦的酰胺鍵斷開,從而開環(huán)與酶形成共價(jià)結(jié)合物,進(jìn)而達(dá)到抑制效果。與他唑巴坦、克拉維酸等經(jīng)典的“自殺式酶抑制劑”不同的是,酶-抑制劑復(fù)合體不發(fā)生水解,經(jīng)環(huán)合形成內(nèi)酰胺環(huán)可又形成阿維巴坦,因此具有長(zhǎng)效抑酶作用[20]。
3 產(chǎn)KPC酶肺炎克雷伯菌對(duì)CZA的耐藥機(jī)制
據(jù)報(bào)道,KPC-Kp對(duì)CZA的耐藥機(jī)制主要有三種,包括blaKPC基因的突變所引起的KPC酶上的氨基酸取代/缺失/插入,KPC酶的表達(dá)量增加和細(xì)菌細(xì)胞膜孔蛋白突變。
3.1 blaKPC基因的突變
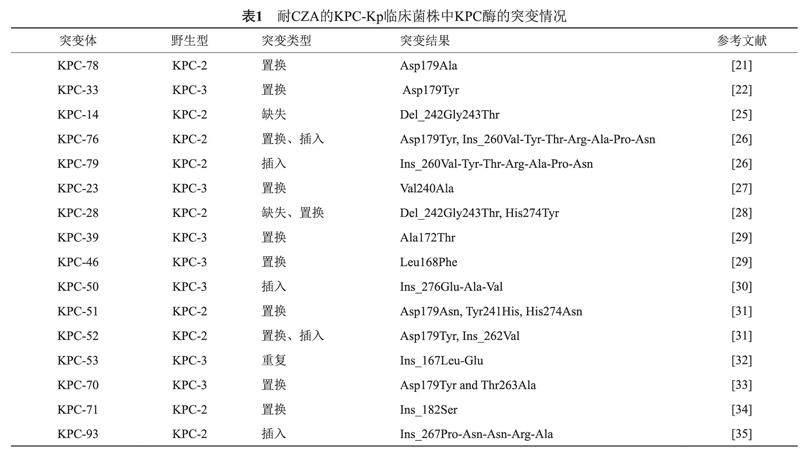
blaKPC基因由879個(gè)堿基組成,當(dāng)基因發(fā)生突變時(shí),會(huì)引起肺炎克雷伯菌所表達(dá)的KPC酶發(fā)生變化,當(dāng)blaKPC基因出現(xiàn)堿基置換時(shí),其結(jié)果對(duì)應(yīng)的是氨基酸的單取代或多取代;當(dāng)blaKPC基因出現(xiàn)多個(gè)堿基缺失和插入時(shí),其結(jié)果對(duì)應(yīng)的是KPC酶的氨基酸缺失或者插入,值得注意的是,臨床上所出現(xiàn)的blaKPC基因變體所插入或者缺失的堿基數(shù)均是3的倍數(shù),所以并未出現(xiàn)移碼突變而導(dǎo)致KPC酶的結(jié)構(gòu)大幅改變。近年來(lái),攜帶blaKPC基因亞型的肺炎克雷伯菌在世界范圍內(nèi)迅速出現(xiàn),自2020年以來(lái)報(bào)告的新型KPC酶變體數(shù)量超過(guò)了前17年的總和。迄今為止,NCBI參考基因目錄共收錄了123個(gè)KPC酶變體,其中有16個(gè)變體來(lái)自臨床報(bào)道的CZA耐藥肺炎克雷伯菌,于表1中列出。
總體而言,導(dǎo)致肺炎克雷伯菌對(duì)CZA耐藥產(chǎn)生的blaKPC基因突變以置換為主,表現(xiàn)為KPC酶的氨基酸取代。其中,以第179位的天冬氨酸被其余氨基酸取代最為常見。研究顯示在一株分型為ST258的臨床肺炎克雷伯菌中發(fā)現(xiàn)了KPC-3變體Asp179Tyr,命名為KPC-33;另有研究報(bào)道在一株分型為ST11的臨床菌株中發(fā)現(xiàn)了KPC-2變體Asp179Ala,命名為KPC-78;這些菌株對(duì)CZA的敏感性均下降,其對(duì)CZA的最小抑菌濃度(Minimum inhibitory concentration, MIC)與親本菌株相比分別上升了64倍和16倍[17-18]。研究顯示通過(guò)定點(diǎn)飽和突變技術(shù)獲得了19種不同的Asp179變異體,并使用質(zhì)譜分析和比較酶促動(dòng)力學(xué)參數(shù)確定了一種特別值得關(guān)注的變異體—Asp179Asn,其可以幫助肺炎克雷伯菌對(duì)多種β-內(nèi)酰胺類藥物和β-內(nèi)酰胺類抑制劑的組合產(chǎn)生抗性[23]。Alsenani等[24]在分析了Asp179N和Asp179Tyr的晶體結(jié)構(gòu)之后,發(fā)現(xiàn)Asp179Asn和Asp179Tyr的存在破壞了Ω環(huán)上鹽橋的穩(wěn)定性,從而使得KPC酶更容易與頭孢他啶結(jié)合,從而使得肺炎克雷伯菌對(duì)CZA耐藥。同樣的機(jī)理可以用以解釋KPC-39(Ala172Thr)和KPC-46(Leu168Phe)變體所引起的CZA抗性,因?yàn)樗鼈兊耐蛔兾稽c(diǎn)也處于KPC酶的Ω環(huán)上(Arg164 – Asp179),其上的突變會(huì)導(dǎo)致分子間的氫鍵作用減弱,從而使得Ω環(huán)結(jié)構(gòu)變得不穩(wěn)定而易于頭孢他啶結(jié)合;同時(shí),有研究報(bào)道Ω環(huán)上鹽橋被破壞后,會(huì)使得KPC酶與阿維巴坦的親和力降低,從而使得阿維巴坦不能有效抑制KPC酶。
除氨基酸取代外,由于KPC酶上出現(xiàn)氨基酸插入和缺失等突變而引起的CZA耐藥在近幾年也時(shí)有發(fā)生。Niu等[25]在紐約分離的一株臨床肺炎克雷伯菌中,存在 KPC-14變體,該變體是由KPC-2缺失第242位甘氨酸和第243位蘇氨酸而形成;在將blaKPC-2和blaKPC-14基因克隆并構(gòu)建到pET-28質(zhì)粒后,并比較了KPC-2和KPC-14的酶促動(dòng)力學(xué)參數(shù),發(fā)現(xiàn)了 KPC-14對(duì)于頭孢他啶的親和力明顯上升,因此可以介導(dǎo)CZA的耐藥。研究報(bào)道在瑞士分離的一株臨床肺炎克雷伯菌中,發(fā)現(xiàn)了KPC-3的變體,命名為KPC-50,其在276至277位插入了谷氨酸、丙氨酸和纈氨酸等三個(gè)氨基酸;Shi等[26]也確定了新變體KPC-76和KPC-79,它們都在KPC-2的第261位至262位中插入了纈氨酸、酪氨酸、蘇氨酸、精氨酸、丙氨酸、脯氨酸和天冬酰胺七個(gè)氨基酸,KPC-76和KPC-79的區(qū)別在于前者多一個(gè)Asp179Tyr突變。耐人尋味的是,Shi等提到了在治療KPC-kp感染中KPC酶多次突變的現(xiàn)象,在治療9 d后,KPC-2突變成了KPC-79,隨后又在CZA治療21 d后突變成了KPC-76,但后期使用美羅培南進(jìn)行治療后又恢復(fù)突變成KPC-2,這說(shuō)明了KPC酶具有高度的靈活性以幫助宿主適應(yīng)環(huán)境的威脅。
值得注意的是,有多項(xiàng)研究報(bào)道了攜帶KPC變體的肺炎克雷伯菌對(duì)除頭孢他啶外其他β-內(nèi)酰胺類藥物的敏感性逆轉(zhuǎn),這一方面提示了美羅培南等β-內(nèi)酰胺類藥物可以作為治療CZA耐藥肺炎克雷伯菌感染的備選方案,另一方面卻增加了快速檢測(cè)KPC-Kp的難度,因?yàn)槠鋵?duì)β-內(nèi)酰胺類藥物的MIC值較低而無(wú)法被篩選。
3.2 膜孔蛋白突變和KPC酶高表達(dá)
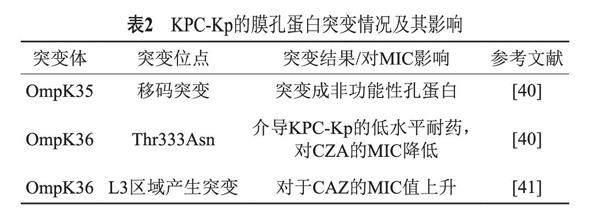
KPC-Kp的膜孔蛋白突變和KPC酶的高表達(dá)均可導(dǎo)致細(xì)菌對(duì)CZA耐藥,并且在某些時(shí)候這兩種因素是偶聯(lián)在一起的。膜孔蛋白發(fā)揮著重要的生理作用,可以促進(jìn)小分子親水性溶質(zhì)的流入,并促進(jìn)有毒物質(zhì)通過(guò)原本不透水的革蘭陰性外膜的流出。與CZA耐藥相關(guān)的細(xì)菌膜孔蛋白主要包括OmpC和OmpF,其中尤以O(shè)mpK35和OmpK36最受關(guān)注,也是近年來(lái)研究的熱點(diǎn)[32-33]。OmpK35和OmpK36的整體結(jié)構(gòu)顯示出與其他一般孔蛋白相似的特征,即由16個(gè)β-折疊片組成的三聚體結(jié)構(gòu)。孔蛋白中的兩個(gè)重要結(jié)構(gòu)成分是細(xì)胞外L3和L4區(qū)域[38](Major nonspecific porins are matrix porin,OmpF)。Humphries等[39]將CZA耐藥株KP1244和敏感株KP1245比較分析后,發(fā)現(xiàn)前者的ompK35基因由于發(fā)生了移碼突變,導(dǎo)致OmpK35不能正常表達(dá)而變成了一個(gè)非功能性孔蛋白。此外,前者與后者相比在ompK36中存在一個(gè)突變—Thr333Asn;進(jìn)一步研究發(fā)現(xiàn),將野生型ompK36轉(zhuǎn)移到KP1244后,其對(duì)CZA的MIC值降低了,說(shuō)明Thr333Asn突變可以介導(dǎo)KPC-Kp的低水平耐藥。Castanheira等[40]也證明了在ompK36的L3區(qū)域產(chǎn)生的突變可以導(dǎo)致細(xì)菌對(duì)于頭孢他啶的MIC值上升(表2)。KPC酶的高表達(dá)也是近年來(lái)導(dǎo)致CZA耐藥的原因之一。2017年有報(bào)道通過(guò)熒光定量PCR技術(shù)發(fā)現(xiàn)一株耐CZA的KPC-3陽(yáng)性肺炎克雷伯菌中blaKPC基因的表達(dá)量是敏感菌株的3.8倍[34-36]。Zhang等[42]對(duì)12株CZA耐藥的KPC-Kp研究后發(fā)現(xiàn),耐藥菌株可通過(guò)增加blaKPC-2所在質(zhì)粒的拷貝數(shù)從而介導(dǎo)KPC酶的高表達(dá);值得注意的是,在將酶抑制劑阿維巴坦的濃度提升之后,大部分耐藥菌株可以恢復(fù)對(duì)CZA的敏感性,說(shuō)明耐藥菌株中KPC酶量的增加使得阿維巴坦的量相對(duì)不足而導(dǎo)致耐藥。Coppi等[44]也發(fā)現(xiàn)KPC-Kp通過(guò)多拷貝攜帶blaKPC-3的轉(zhuǎn)座子Tn4401a從而使得blaKPC-3基因高表達(dá),同時(shí)研究也發(fā)現(xiàn)blaKPC-3基因高表達(dá)跟外膜蛋白的結(jié)構(gòu)變化相關(guān),blaKPC-3基因高表達(dá)使得細(xì)菌的膜孔蛋白的通透性降低,從而導(dǎo)致對(duì)CZA的耐藥性升高。但是,在Sun等[44]的研究中,blaKPC基因的突變和其高表達(dá)共同導(dǎo)致了KPC-Kp對(duì)CZA的抗性增加,然而其膜孔蛋白的突變對(duì)CZA的易感性并無(wú)影響。
4 小結(jié)和展望
綜上所述,KPC陽(yáng)性肺炎克雷伯菌對(duì) CZA產(chǎn)生耐藥性主要是由于blaKPC基因的突變、細(xì)菌細(xì)胞膜孔蛋白突變和KPC酶的表達(dá)量增加,這三種機(jī)制有可能同時(shí)出現(xiàn)而導(dǎo)致KPC-Kp對(duì)CZA的高水平耐藥。針對(duì)KPC-Kp的感染,在使用CZA進(jìn)行治療的同時(shí),臨床醫(yī)生應(yīng)當(dāng)密切關(guān)注KPC-Kp對(duì)美羅培南等碳青霉烯類抗生素的敏感性變化,依據(jù)實(shí)際情況進(jìn)行聯(lián)合用藥和輪換用藥。
參 考 文 獻(xiàn)
WHO. Antimicrobial resistance. Global report on surveillance[J]. World Health Organization, 2014, 61(3): 12-28.
Lai Y C, Lu M C, Hsueh P R. Hypervirulence and carbapenem resistance: two distinct evolutionary directions that led high-risk Klebsiella pneumoniae clones to epidemic success[J]. Expert Rev Mol Diagn, 2019, 19(9): 825-837.
Hennequin C, Robin F. Correlation between antimicrobial resistance and virulence in Klebsiella pneumoniae[J]. Eur J Clin Microbiol Infect Dis, 2016, 35(3): 333-341.
Navon-Venezia S, Kondratyeva K, Carattoli A. Klebsiella pneumoniae: a major worldwide source and shuttle for antibiotic resistance[J].FEMS Microbiol Rev, 2017, 41(3): 252-275.
Patel G, Huprikar S, Factor S H, et al. Outcomes of carbapenem-resistant Klebsiella pneumoniae infection and the impact of antimicrobial and adjunctive therapies [J]. Cambridge University Press (CUP), 2008, 29(12): 1099-1106.
Tumbarello M, Viale P, Viscoli C, et al. Predictors of mortality in bloodstream infections caused by Klebsiella pneumoniae carbapenemase-producing K. pneumoniae: importance of combination therapy [J]. Clin Infect Dis, 2012, 55:943-950.
Hu F, Wang M, Zhu D, et al. CHINET efforts to control antimicrobial resistance in China[J].J Glob Antimicrob Resist, 2020, 21: 76-77.
Liao W, Liu Y, Zhang W. Virulence evolution, molecular mechanisms of resistance and prevalence of ST11 carbapenem-resistant Klebsiella pneumoniae in China: A review over the last 10 years[J]. J Glob Antimicrob Resist, 2020, 23: 174-180.
Qi Y, Wei Z, Ji S, et al. ST11, the dominant clone of KPC-producing Klebsiella pneumoniae in China[J].? J Antimicrob Chemother, 2011, 66(2): 307-312.
Hernández-García M, Castillo-Polo J A, Cordero D G, et al. Impact of ceftazidime-avibactam treatment in the emergence of novel KPC variants in the ST307-Klebsiella pneumoniae high-risk clone and consequences for their routine detection[J]. J Clin Microbiol, 2022, 60(3):e01429.
Zhang Y, Jin L, Ouyang P, et al. Evolution of hypervirulence in carbapenem-resistant Klebsiella pneumoniae in China: a multicentre, molecular epidemiological analysis[J]. The Journal of antimicrobial chemotherapy[J]. J Antimicrob Chemother, 2020, 75(2): 327-336.
Xu L, Sun X, Ma X. Systematic review and meta-analysis of mortality of patients infected with carbapenem-resistant Klebsiella pneumoniae[J]. Ann Clin Microbiol Antimicrob, 2017, 16(1):18.
Zhang Y, Wang Q, Yin Y, et al. Epidemiology of carbapenem-resistant Enterobacteriaceae infections: report from the China CRE network[J]. Antimicrob Agents Chemother, 2018, 62(2):e01882-17.
Mendes R E, Castanheira M, Gasink L, et al. β-lactamase characterization of Gram-negative pathogens recovered from patients enrolled in the phase 2 trials for ceftazidime-avibactam: Clinical efficacies analyzed against subsets of molecularly characterized isolates[J]. Antimicrob Agents Chemother, 2015, 60(3): 1328-1335.
van Duin D, Bonomo R A. Ceftazidime/avibactam and ceftolozane/tazobactam: second-generation β-lactam/β-lactamase inhibitor combinations[J]. Clin Infect Dis, 2016, 63(2): 234-241.
Stone G G, Bradford P A, Newell P, et al. In Vitro activity of ceftazidime-avibactam against Isolates in a phase 3 open-label clinical trial for complicated intra-abdominal and urinary tract infections caused by ceftazidime-nonsusceptible Gram-negative pathogens[J]. Antimicrob Agents Chemother, 2017, 61(2):e01820-16.
Li D, Liao W, Huang H H, et al. Emergence of hypervirulent ceftazidime/ avibactam-resistant Klebsiella pneumoniae isolates in a Chinese tertiary hospital[J]. Infect Drug Resist, 2020, 13: 2673-2680.
Compain F, Arthur M. Impaired inhibition by avibactam and resistance to the ceftazidime-avibactam combination due to the D179Y substitution in the KPC-2 β-lactamase[J]. Antimicrob Agents Ch, 2017, 61(7):e00451-17.
Humphries R M, Yang S, Hemarajata P, et al. First report of ceftazidime-avibactam resistance in a KPC-3-expressing Klebsiella pneumoniae isolate[J]. Antimicrob Agents Chemother, 2015, 59(10): 6605-6607.
Matesanz M, Mensa J. Ceftazidime-avibactam revista espanola de quimioterapia : publicacion oficial de la Sociedad Espanola de Quimioterapia[J]. Rev Esp Quimioter, 2021, 34(Suppl1): 38-40.
Liao Q, Deng J, Feng Y, et al. Emergence of ceftazidime-avibactam resistance due to a novel blaKPC-2 mutation during treatment of carbapenem-resistant Klebsiella pneumoniae infections[J]. J Infect Public Health, 2022, 15(5): 545-549.
Shields R K, Chen L, Cheng S, et al. Emergence of ceftazidime-avibactam resistance due to plasmid-borne blaKPC-3 mutations during treatment of carbapenem-resistant Klebsiella pneumoniae infections[J]. Antimicrob Agents Chemother, 2017, 61(3):e02097-16.
Barnes M D, Winkler M L, Taracila M A, et al. Klebsiella pneumoniae carbapenemase-2 (KPC-2), ubstitutions at ambler position Asp179, and resistance to ceftazidime-avibactam: Unique antibiotic-resistant phenotypes emerge from β-lactamase protein engineering[J]. mBio, 2017, 8(5):e00528-17.
Alsenani T A, Viviani S L, Kumar V, et al. Structural characterization of the D179N and D179Y variants of KPC-2 β-lactamase: Ω-loop destabilization as a mechanism of resistance to ceftazidime-avibactam[J]. Antimicrob Agents Chemother, 2022, 66(4):e0241421.
Niu S, Chavda K D, Wei J, et al. A ceftazidime-avibactam-resistant and carbapenem-susceptible Klebsiella pneumoniae strain harboring blaKPC-14 isolated in New York city[J]. mSphere, 2020, 5(4):e00775-20.
Shi Q, Han R, Guo Y, et al. Multiple novel ceftazidime-avibactam-resistant variants of blaKPC-2-Positive Klebsiella pneumoniae in two patients[J]. Microbiology spectrum, Microbiol Spectr, 2022, 10(3).
Galani I, Antoniadou A, Karaiskos I, et al. Genomic characterization of a KPC-23-producing Klebsiella pneumoniae ST258 clinical isolate resistant to ceftazidime-avibactam[J]. Clin Microbiol Infect, 2019, 25(6): 763.e5-763.e8.
Oueslati S, Iorga B I, Tlili L, et al. Unravelling ceftazidime/avibactam resistance of KPC-28, a KPC-2 variant lacking carbapenemase activity[J]. J Antimicrob Chemother, 2019, 74(8): 2239-2246.
Jousset A B, Oueslati S, Emeraud C, et al. KPC-39-mediated resistance to ceftazidime-avibactam in a Klebsiella pneumoniae ST307 clinical isolate[J]. Antimicrob Agents Chemother, 2021, 65(12):e0116021.
Poirel L, Vuillemin X, Juhas M, et al. KPC-50 confers resistance to ceftazidime-avibactam associated with reduced carbapenemase activity[J].Antimicrob Agents Chemother, 2020, 64(8):e00321-20.
Sun L, Chen W, Li H, et al. Phenotypic and genotypic analysis of KPC-51 and KPC-52, two novel KPC-2 variants conferring resistance to ceftazidime/avibactam in the KPC-producing Klebsiella pneumoniae ST11 clone background[J].J Antimicrob Chemother, 2020, 75(10): 3072-3074.
Di Pilato V, Aiezza N, Viaggi V, et al. KPC-53, a KPC-3 variant of clinical origin associated with reduced susceptibility to ceftazidime-avibactam[J]. Antimicrob Agents Chemother, 2020, 65(1):e01429-20.
Mendes G, Ramalho J F, Bruschy-Fonseca A, et al. First description of ceftazidime/avibactam resistance in a ST13 KPC-70-producing Klebsiella pneumoniae strain from portugal[J]. Antibiotics (Basel), 2022, 11(2):167.
Li X, Ke H, Wu W, et al. Molecular mechanisms driving the in vivo development of KPC-71-mediated resistance to ceftazidime-avibactam during treatment of carbapenem-resistant Klebsiella pneumoniae infections[J]. mSphere, 2021, 6(6):e0085921.
Wu Y, Yang X, Liu C, et al. Identification of a KPC variant conferring resistance to ceftazidime-avibactam from ST11 carbapenem-resistant Klebsiella pneumoniae strains[J]. Microbiol Spectr, 2022, 10(2):e0265521.
Shields R K, Clancy C J, Hao B, et al. Effects of Klebsiella pneumoniae carbapenemase subtypes, extended-spectrum β-lactamases, and porin mutations on the in vitro activity of ceftazidime-avibactam against carbapenem-resistant K. pneumoniae[J]. Antimicrob Agents Chemother, 2015, 59(9): 5793-5797.
Pagès J M, Peslier S, Keating T A, et al. Role of the outer membrane and porins in susceptibility of β-lactamase-producing enterobacteriaceae to ceftazidime-avibactam[J]. Antimicrob Agents Chemother, 2015, 60(3): 1349-1359.
Dutzler R, Rummel G, Alberti S, et al. Crystal structure and functional characterization of OmpK36, the osmoporin of Klebsiella pneumoniae[J]. Structure, 1999, 7(4): 425-434.
Humphries R M, Hemarajata P. Resistance to ceftazidime-avibactam in Klebsiella pneumoniae due to porin mutations and the increased expression of KPC-3[J]. Antimicrob Agents Chemother, 2017, 61(6):e00537-17.
Castanheira M, Doyle T B, Hubler C, et al. Ceftazidime-avibactam activity against a challenge set of carbapenem-resistant Enterobacterales: Ompk36 L3 alterations and β-lactamases with ceftazidime hydrolytic activity lead to elevated MIC values[J]. Int J Antimicrob Agents, 2020, 56(1):106011.
Humphries R M, Yang S, Hemarajata P, et al. First report of ceftazidime-avibactam resistance in a KPC-3-expressing Klebsiella pneumoniae isolate[J]. Antimicrob Agents Chemother, 2015, 59(10): 6605-6607.
Zhang P, Shi Q, Hu H, et al. Emergence of ceftazidime/avibactam resistance in carbapenem-resistant Klebsiella pneumoniae in China[J]. Clin Microbiol Infect, 2020, 26(1): 124.e1-124.e4.
Coppi M, Di Pilato V, Monaco F, et al. Ceftazidime-avibactam resistance associated with increased blaKPC-3 gene copy number mediated by pKpQIL plasmid derivatives in sequence type 258 Klebsiella pneumoniae[J]. Antimicrob Agents Chemother, 2020, 64(4):e01816-19.
Sun L, Li H, Wang Q, et al. Increased gene expression and copy number of mutated blaKPC lead to high-level ceftazidime/avibactam resistance in Klebsiella pneumoniae[J]. BMC Microbiol, 2021, 21(1):230.
收稿日期:2022-11-06
作者簡(jiǎn)介: 鄭媚,碩士研究生,主要從事細(xì)菌耐藥性研究。
*通訊作者:于洋,副教授,主要從事細(xì)菌耐藥性以及藥動(dòng)藥代等研究。
猜你喜歡
中國(guó)現(xiàn)代醫(yī)生(2017年4期)2017-04-26 09:34:35
中國(guó)醫(yī)藥科學(xué)(2016年20期)2017-03-15 19:46:37
農(nóng)業(yè)與技術(shù)(2016年20期)2016-12-08 21:34:56
體育教學(xué)(2016年10期)2016-12-01 00:16:53
考試周刊(2016年76期)2016-10-09 09:47:35
江蘇農(nóng)業(yè)科學(xué)(2016年4期)2016-06-14 01:38:13
環(huán)球時(shí)報(bào)(2015-06-25)2015-06-25 19:30:56
安徽農(nóng)學(xué)通報(bào)(2015年10期)2015-06-15 02:32:43
湖北農(nóng)業(yè)科學(xué)(2014年22期)2015-01-20 11:21:07
江蘇農(nóng)業(yè)科學(xué)(2014年11期)2015-01-15 12:13:21
