針闊混交林生態系統特征及生態效益分析
2021-01-13 20:21:50潘婷雷云申玲芝朱曉春徐明張健
山地農業生物學報 2021年5期
關鍵詞:生態系統
潘婷 雷云 申玲芝 朱曉春 徐明 張健
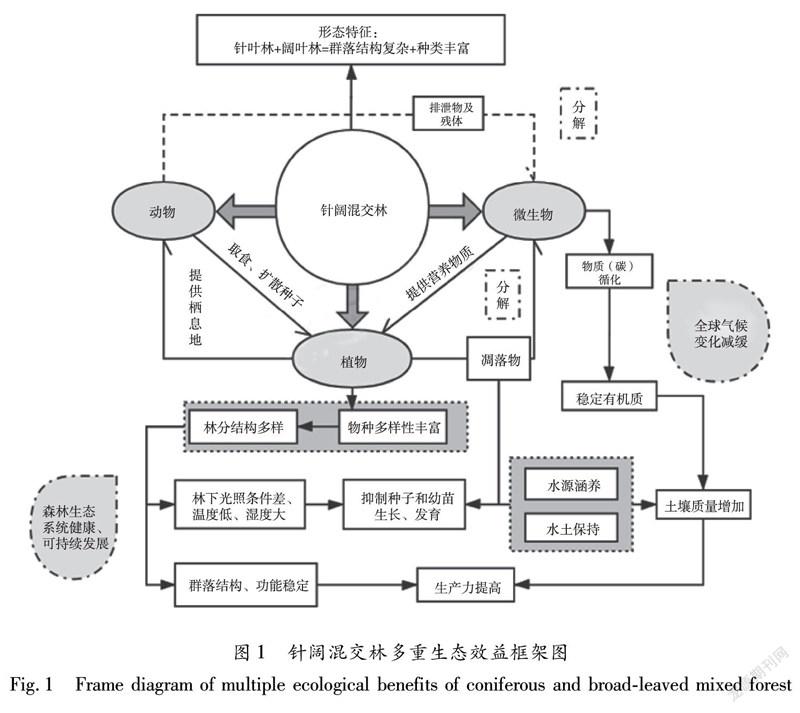
摘 要:針闊混交林是一種常見、可提供多重生態服務功能的森林類型,相對于人工純林在生態效益、防護效益以及經濟效益等方面具有明顯優勢。本文介紹了針闊混交林生態系統的生物多樣性、生產力、空間結構、土壤養分、凋落物等方面特征,對其生態效益進行分析,以期為針闊混交林生態系統的科學管理與利用提供科學依據。
關鍵詞:針闊混交林;生態系統;生態效益
中圖分類號:S718.5? 文獻標識碼:A
文章編號:1008-0457(2021)05-0040-08? ? 國際DOI編碼:10.15958/j.cnki.sdnyswxb.2021.05.007
Analysis of Ecosystem Characteristics and Ecological Benefit of Coniferous and Broad-leaved Mixed Forest
PAN Ting,LEI Yun,SHEN Lingzhi,ZHU Xiaochun,XU Ming,ZHANG Jian*
(College of Life Sciences,Guizhou University,Guiyang,Guizhou 550025,China)
Abstract:Coniferous and broad-leaved mixed forest is a common forest type that can provide multiple ecological services.Compared with pure artificial forest,it has obvious advantages in ecological,protection and economic benefits.This paper introduced the characteristics of biodiversity,productivity,spatial structure,soil nutrients and litter of coniferous and broad-leaved mixed forest ecosystem,and analyzed its ecological benefits,in order to provide scientific basis for scientific management and utilization of coniferous and broad-leaved mixed forest ecosystem.
Keywords:coniferous and broad-leaved mixed forest;ecosystem;ecological benefits
基金項目:國家自然科學基金(31960234);貴州省科技支撐計劃項目(黔科合支撐[2018]2773);大學生創新創業訓練計劃項目(S202010657083)
森林作為地球上最重要的陸地生態系統類型,有“地球之肺”的美譽,擁有著豐富的生物多樣性和復雜的群落結構與功能,承載著陸地上70%的物種,對全球氣候的調節與穩定有著重大意義[1]。森林生態系統(Forest Ecosystem)是森林中生物(以喬木為主)與其環境、生物與生物之間相互作用、相互依存的統一整體,按樹種組成可分為純林和混交林。混交林一般由兩個或多個優勢喬木樹種或不同生活型的喬木所組成的森林。針闊混交林(Coniferous and broad-leaved mixed forests)是由針葉樹種與闊葉樹種為優勢種共存的天然或人工混交林,是介于針葉林和闊葉林之間的過渡性植被類型。受區域氣候影響,針闊混交林的種類組成在不同地區存在明顯的差別。全球的溫帶針闊混交林主要分布于北半球北緯40°~60°之間的歐洲西緣、北美洲東緣和亞洲東緣,其物種多樣性豐富,由櫟樹、椴樹等夏綠闊葉樹種和云杉、冷杉、松樹等針葉樹種混合組成,在群落演替過程中具有較高的研究價值。我國針闊混交林一般分為溫帶針葉、落葉闊葉混交林和亞熱帶山地針葉、常綠闊葉、落葉闊葉混交林兩種類型,主要分布于東北地區(如大、小興安嶺和長白山山地)[2-3]和南方山地地區(如西南山地亞高山和中山的林區)。
我國是世界上人工林建設面積最大的國家[4],但仍以純林為主,如馬尾松純林、樟子松純林等,人工純林存在病蟲害多發、地力衰退嚴重、土壤環境惡化、易受外界環境波動的影響、森林更新困難等生態問題,而且還容易導致火災的發生和蔓延[4-6]。天然針闊混交林在涵養水源,水土保持能力,提高生長率,提供多樣化林產品,減少病蟲害發生幾率,防止火災蔓延等方面發揮了重要的生態作用,在構建新型高生態、經濟、社會效益的多功能森林生態體系方面也占有重要戰略地位[2]。因此,有必要加強針闊混交林生態系統的基礎理論與生態環境效益評價等方面的深入研究,以期為針闊混交林的科學管理與利用提供參考。
1 針闊混交林生態系統主要特征
針闊混交林植物區系的地理成分較為復雜,植物生活型譜以高位芽植物為主,其次為地面芽植物,含有一定比例的地下芽和地上芽植物,一年生植物種類極少[7]。實踐證明,混交林能夠有效地增加林地生物物種的多樣性,提高林地養分水平和生產力,從根本上改善林分的生態環境[8]。
1.1 林分結構、多樣性與生產力相互關系
物種多樣性和生產力是森林生態系統的重要特征,而林分結構可以強烈地影響森林的生長和其他生態過程,如水的平衡、碳分配、營養物質的循環和光的動力學[9]。不同于多樣性的間接作用,林分結構能直接影響森林生產力。植物種間互作將有利于增加在自然干擾、脅迫或捕食等條件下難于生存的植物種類,從而促進森林群落物種多樣性增加[10]。“生態位互補假說”認為不同物種由于生態位差異會對資源進行更充分的利用,從而可提高森林生產力[11]。針闊混交林中不同林層林分結構對生產力的作用方向不同,林下層的物種多樣性較林冠層的物種多樣性對生產力影響弱,這主要是由于樹木生長發育和環境異質性的綜合作用[12]。
植物個體的生長、死亡和更新等動態將直接影響群落空間結構,良好的林分空間結構是判斷群落能否維持穩定發展的重要前提。混交度是描述混交林中不同樹種之間隔離程度的重要空間結構指數[13]。樹種混交度是群落發育過程中種群與環境、種群與種群之間長期相互作用的結果。混交林中樹種之間的隔離程度由樹種類型及其不同樹種數量和空間分布共同決定。一般群落的樹種類型越多,其混交度也越大。群落混交程度越高反映出其群落結構越穩定。周紅敏等[14]在鳳陽山自然保護區針闊混交林空間結構研究中指出林木混交度較強,物種多樣性高,林分空間結構相對穩定。朱軍等[15]發現紫金山針闊混交林的混交度對林下植物多樣性具有重要影響。混交林相對純林的空間結構多樣性較復雜;混交林的物種多樣性、群落更新潛力和群落穩定性均高于純林。混交林中的樹種混交比例會改變森林內物種豐富度和多樣性、種群更新能力、群落生物量等,從而影響森林群落的穩定性[16]。
森林空間結構是森林生長過程的重要驅動因子,對森林的功能表達和發展趨勢具有關鍵性作用。物種的種類與空間分布格局決定群落空間結構,物種間不同空間關系塑造了群落結構的特異性,良好的空間結構是群落能夠維持穩定發展的前提條件。在一個群落中,優勢種作為群落演替的內在驅動力,最能體現一個群落內部的空間結構和格局的形成過程。林分空間結構決定林木間的競爭優勢及其空間生態位,在很大程度上決定林分的穩定性、發展的可能性。有學者提出從林木空間分布格局、樹種混交度和樹木競爭等方面反映林分內部的結構變化[17]。林木空間分布格局包括聚集分布、隨機分布和均勻分布等形式。李建等[18]研究發現長白山次生針闊混交林群落的水平格局為隨機分布,大小分化程度為中庸分布且樹種混交度較強時,群落具有良好且較穩定的空間結構。
群落結構受生境異質性、植物繁殖體傳播范圍和種間相互作用等因素影響且隨其研究尺度的變化而變化。不同的研究尺度可能會得出不同的分布類型。在較小的尺度范圍內可能受到種內或種間競爭、種子擴散限制等因素的影響,而在大尺度范圍內可能受自身的生物學特性、種群分布區的異質性和外界無機環境等因素的影響[19]。近年來,許多學者對我國長白山地區針闊混交林空間格局進行了大量研究,為我國針闊混交林空間格局的進一步研究奠定了基礎[20-24]。
1.2 土壤性質
林地土壤肥力主要靠林木養分的歸還量和養分循環速度來維持和提高,并可直接影響植物生長,在植物體構成和生理代謝方面發揮著重要作用。混交林中植被的種類多數量大,對土壤涵養水源和累積有機質有巨大作用,明顯改善了土壤的通透性,降低了土壤的緊實程度和容重,特別是表層土壤最為明顯[25]。混交林中土壤最上層有大量的凋落物,它們分解后成為表層土,大大改善了表層土的物理性狀,增加了土壤有機質和團粒結構,提高了土壤孔隙度,減小了土壤容重。在混交林中,由于增加了闊葉樹種,加速了枯枝落葉的分解速率,各營養元素的釋放量得到提高[26],故與純林相比,所有混交林中同一土層中的有機質、全氮、水解氮、全磷、速效磷、全鉀、速效鉀的含量都有不同層度的增加。有更厚的枯枝落葉層和腐殖質層,較純林地其土壤質地松散、結構優異、吸持水的能力強、透水性大[27]。土壤的物理性狀尤其是土壤類型、孔隙度和結構等與持水能力、保水能力及滲水能力等方面直接相關,影響著林地植物對土壤養分的吸收和保持。針闊混交林的土壤理化性質隨演替過程發生變化,毛管孔隙度、速效鉀和全磷含量是影響不同演替階段土壤質量的主要因子;經過不同演替階段,表層土壤理化性質質量的綜合得分表現為中齡林 < 成熟林 < 近熟林 < 老齡林;隨著演替進行,針闊混交林的表層土壤理化性質基本呈現質量提高趨勢,到老齡林時期達到最佳[28]。
1.3 凋落物
森林的凋落物在森林資源保護、可持續利用方面發揮著重要作用,且在維持土壤肥力,保證植物再生長養分的可循環利用性,促進森林生態系統正常的物質循環和養分平衡方面起著重要的作用[29]。凋落物是林木養分歸還的主要途徑,凋落物積累量和分解速率決定著林木養分歸還量的大小[30];一般而言,凋落物分解速率與N、P含量成正比。凋落物量是一個動態值,它受氣候、地形、海拔、土壤和植被類型等多因素影響。從熱帶到亞熱帶和溫帶,凋落物現存量隨著緯度的增高而增加的結論基本一致。凋落物層具有調節和阻滯地表徑流的生態水文功能,如緩沖雨水的動能,避免土壤結構受雨水濺擊而破壞[31]。針對我國東北地區地帶性頂級植物群落——針闊混交林的研究表明,其凋落物量平均為4.49 t/hm2,其中針、闊葉的凋落物量分別占全年的23%和60%左右[32]。凋落物的水源涵養功能由凋落物量和持水能力共同決定;一般情況下,凋落物的現存量越多,持水能力越強,其水源涵養功能越好[33]。針闊混交林的凋落量較大,且在不同森林類型中營養元素的儲量一般表現為:未分解層 <半分解層 < 完全分解層[34]。總之,凋落物的分解對森林生態系統的結構和功能有著不可忽視的影響。
1.4 針闊混交林相關的生態機制
1.4.1 物種更新與維持機制
幼苗和幼樹是植物群落更新的關鍵,是群落維持穩定的決定因素,且直接影響種群繁衍[35]。幼苗作為林木更新過程中最敏感脆弱的環節,其存活狀況直接影響森林群落的物種組成和生態系統的穩定性[36]。針闊混交林由于林下光照條件差、灌木、草本層豐富,對幼苗和幼樹形成了競爭抑制作用。再加上混交林下腐殖質及枯枝落葉層較厚,大大降低了種子接觸土壤萌發的幾率。已有研究表明,林下幼苗種類及物種豐富度隨著采伐強度的增加而增加。首先,適度采伐可改善林下微環境(如光照和土壤溫度)、優化林分結構、調整林分密度,使林下光照增多,利于耐陰樹種的生長,影響著林下植被多樣性,進而影響林下植被的更新和生長。其次,采伐也可能使森林樹種的產子量增加,充足的光照和適宜的土壤溫度促進種子萌發,提高物種多樣性[37-38]。
物種多樣性維持及其空間尺度效應是生態學研究的重點問題[39],不同空間尺度下生態學過程可能存在差異[40]。在不同尺度上植物群落物種多樣性的發生和維持主要通過物種生物學和生態學特性的差異和生境異質性得以實現[41]。基于單物種—面積模型,可將物種對鄰體多樣性格局的影響區分為促進種、中性種和抑制種。生物多樣性促進種在群落多樣性格局組建過程中起主導作用,中性種所代表的隨機過程在群落組建過程中所發揮的作用也不容忽視。同時,在中度和高度脅迫的環境中,植物間正向互作通過擴展現實生態位,可以提高群落生物多樣性[42]。
1.4.2 競爭效應
競爭指2個或多個植物體在同一環境資源和能量的爭奪中所發生的相互作用;研究樹種的種內種間競爭特性對改善森林結構、提高森林景觀品質具有重要作用;森林物種競爭關系研究是森林生態學的重要研究領域之一。辛營營等[43]通過對杭州青山湖地區針闊混交林優勢種的數量研究發現,優勢樹種的種內競爭大于種間競爭,說明優勢樹種呈現出一定的聚集分布狀態,競爭強度與胸徑大小之間存在著冪函數關系,但當胸徑達到一定程度即樹種相對成熟穩定時兩者之間的關系不再明顯,認為這為針闊混交林的可持續經營實踐中“間密留稀,去弱存強”減低林木間競爭的撫育措施提供了理論支持。杜秀芳等[44]對杭州市臨安區多種森林類型的競爭關系研究表明,該區域森林的競爭主要來源于種內競爭,種內平均競爭順序為:人工針葉林>天然闊葉林>天然針葉林>天然針闊混交林>人工針闊混交林>人工闊葉林;天然針闊混交林種間平均競爭最大,人工針闊混交林次之,針闊混交林的樹種豐富度明顯高于其它。
1.4.3 邊緣效應
邊緣效應指在兩個或多個不同性質的生態系統(或其他系統)交互作用處,由于某些生態因子(可能是物質、能量、信息、時機或地域)或系統屬性的差異和協合作用而引起系統某些組分及行為(如種群密度、生產力和多樣性等)的較大變化。邊緣效應的存在使林內到林外的生態環境產生梯度變化,進而影響動植物分布、群落結構和功能、群落能量流和物質流等[45]。奚為民等[46]通過對縉云山森林林窗及其邊緣效應的研究表明,林窗邊緣區由于邊緣效應的作用而有增大物種多樣性的趨勢,其中針闊混交林中前期林窗的邊緣效應更為顯著;而針闊混交林林窗具有一般意義上的比常綠闊葉林林窗相對較強的邊緣效應強度。林窗邊緣效應規律可以為混交林的種群結構重建以及選擇最佳的林木擇伐面積和強度等提供科學依據。因此,維持一定的林窗邊緣效應強度和邊緣區面積,有利于森林物種多樣性的保持和林木更新。
2 針闊混交林生態效益分析
森林是陸地生態系統的主體,是人類賴以生存的自然環境,具有生態效益、經濟效益和社會效益三大效益。針闊混交林生態系統的多種特征決定了其獨特的生態效益優勢,其物種多樣性優于針葉純林,進而影響著森林生態系統的演替和動態,改變著森林群落的區系組成、結構和遺傳多樣性[47]。
2.1 針闊混交林的生態環境
針闊混交林物種多樣性豐富、林分結構復雜、綜合生產力高、生態系統相對穩定,生態環境良好。通過人工經營的方法,根據適地適樹原則,有目的地將針、闊樹種按一定比例混交種植,在其生長過程中,通過環境選擇和競爭作用能自然形成合理協調的林分組合;同時,與針葉純林相比,在針闊混交林內草本、灌木層植物更為豐富,森林的水平垂直層次結構十分合理[48];即使在混交林被砍伐后,其生態環境破壞較小,易于森林群落的更新。
2.2 針闊混交林的水循環效益
2.2.1 針闊混交林的樹冠截留降雨率
不同林分因樹冠結構特征的不同,其降雨截留作用會存在明顯差異。有研究表明,人工針闊混交林對樹冠截留的功能要強于松葉純林[49]。針闊混交林的多層林冠一般會呈現出垂直互補協調的特點,能夠阻攔、延長和增加雨水流經樹冠的時間。因此,如果是在降雨強度相同且年齡條件相同的基礎上,針闊混交林的林冠截留降雨率要比針葉純林多[48]。林冠對降雨的截留可減少雨水對地面的沖蝕、也影響著土壤中水分的分布及蒸發、蒸騰等過程。林冠對降雨的截留作用揭示了森林與水的關系問題,具有重要的生態水文意義[50]。
2.2.2 針闊混交林的涵養水源和水土保持能力
針闊混交林的枯枝落葉量多,能相互促進分解,吸持雨水能力強,在一定程度上能緩沖雨水對地表的沖擊;再加上植物細根能固定土壤,水土保持能力強。不同森林類型單位面積水源涵養能力一般表現出:常綠闊葉林>針闊混交林>常綠針葉林>落葉闊葉林>灌木林[51]。與針葉純林相比,針闊混交林的枯枝落葉層和腐殖質層相對較厚,具體為:針闊混交林的枯落物總厚度比針葉純林多58%,腐爛物的厚度多121%,持水率多37%[48]。此外,混交林具有數量大、密度高等特點,相對減少了混交林內的光照面積和時間。再加上林內通風效果不佳,水分的蒸發就隨之得到抑制,提高了林內的濕度,改善了混交林的氣候環境[52]。
2.3 針闊混交林的土壤理化性質改善作用
混交林較純林相比,具有多層的樹冠以及茂密的樹葉,其林下物種多樣性較高,凋落物豐富,易于形成松軟易分解的腐殖質,其養分分解能力及轉化速率加快,土壤有機質質量分數高。混交林植物可以有效改善土壤的通氣狀況,使得林地土壤變得疏松多孔,土壤孔隙度增大[53],提高了混交林地的土壤肥力,改善了土壤結構,增強林地的容蓄能力;混交林植物的根系生長迅速,根的吸收功能增強,有效解決純林地力衰退問題,更有利于混交林植物的生長。有學者通過對針闊混交林和人工純松林的土壤性質分析比較發現,人工針闊混交林不僅能提高土壤肥力,較之純林還能吸收更多的二氧化碳,釋放更多的氧氣,從而有效地增加森林的碳匯能力[54]。
3 針闊混交林研究與應用的展望
針闊混交林是世界主要的森林類型之一,具有獨特的森林生態系統結構與功能,發揮著重要的生態效益、防護效益和經濟效益。隨著全球氣候變化和生態環境退化的挑戰,針闊混交林已成為林學、生態學等領域研究熱點之一。國內對針闊混交林的研究主要集中在其分布較為密集的大小興安嶺、長白山等山地,對西南地區也有探究[18,22-24,35,47]。針對針闊混交林的研究與應用是多方面的,包括生物多樣性、生產力、動態變化、空間結構、土壤養分、凋落物、林下種植、近自然經營[55-56]等方面。下面將試圖從造林學、生態學和學科交叉等角度出發,對針闊混交林研究與應用進行展望。
3.1 生態學角度:針闊混交林的生態學機制研究及其生態功能評價? 針闊混交林中物種豐富,有復雜的結構,且林分的郁閉度大,因此造成葉面積指數大,景觀層次及冠層結構相對較好。針闊混交林的枯落物層和土壤層的綜合水源能力優于針葉混交林和杉木林,能改善森林的生態環境,對林分的增產有促進作用,應該加強對針闊混交林的研究保護,提高森林生態系統在涵養水源、保持水土方面的生態功能,充分展現混交林功能效益和潛在的性能[57]。
3.2 造林學角度:構建多重生態功能的針闊混交林? 基于地理環境特征、種間關系、功能性狀、空間格局等多角度的綜合分析針闊混交林群落生態學特征與規律,探究混交林營建的最佳的針、闊混交比例,造林不同階段的生態調控技術,激發混交林的獨特優勢,構建多重生態功能的針闊混交林,創建優良的生態環境,更好滿足人類日益增長森林生態產品的多樣化需求。
3.3 交叉學科角度:針闊混交林的森林康養與林下經濟研究與應用? 森林康養涉及了醫療、旅游、生態、社會和經濟等領域,是林業發展催生的新模式[58],它有助于捕獲碳排放,促進生物多樣性,并在農村和城市周邊景觀中提供娛樂和美學價值[59]。較高的樹種多樣性增加了生態位的數量,從而進一步增加了伴生物種的數量,例如,能為林下植物和動物提供更好的棲息地[60],營造生態、景觀、經濟多效益多功能混交林群落[61]。林下經濟是最近興起的農林復合型產業,可實現農林牧業資源的合理共享和循環協調發展,具有良好的經濟效益和社會效益[62]。目前,林下經濟發展迅速,林下植物和動物都有具有較大發展潛力。同時,還可發展林下文化產業,發展旅游勝地、養生保健、有機食物等。例如:北京市建設林下經濟示范園區發展親子采摘、森林觀光、幼苗種植等。東北地區發展的林間養鹿、種蛙、種參等。云南發展石斛、菌菇和蔬菜等林下產品,取得了顯著的經濟發展和環境保護效果[63]。由于針闊混交林具有物種多樣性豐富、林分結構多樣、形態多樣、腐殖質層厚和生態環境良好等特點,為林下創造了更多的異質光環境,促進了林下動植物生長,對于森林康養和林下經濟的發展提供了優質條件。
針闊混交林對快速恢復森林的生態環境性能,涵養水源,水土保持能力,生長率的提高,產品的多樣化,防護效益的發揮,減少病蟲害發生幾率,防止火災蔓延等方面發揮了巨大作用;同時還能為市場提供多元化、多性能的木制產品,拉動經濟發展。因此,對于針闊混交林進行多方面多層次的研究,對保障森林生態系統持續性和穩定性具有實踐意義,為林分結構化經營、混交林的撫育、林分質量評價和樹種空間多樣性評價等提供了科學依據,同時也為植被恢復與保護、資源利用及生物多樣保護和森林管理提供參考。針闊混交林對水土保持、涵養水源、調節氣候、農業生產和經濟的可持續發展有重要作用,對土壤生產力匱乏地區尤其是喀斯特地區具有良好的生態影響,有較廣闊的發展前景。
參 考 文 獻:
[1]? 蘇巧靈,李安民,袁士云,等.甘肅小隴山暖溫帶針闊混交林物種組成和群落結構[J].應用生態學報,2020,31(10):3305-3312.
[2]? 周秋靜,韓文斌,趙常明,等.神農架天然針闊混交林的物種組成和群落結構[J].生態學雜志,2019,38(1):11-8.
[3]? 張煒琪,陳輝,林文俊,等.武夷山針闊混交林的群落多樣性[J].森林與環境學報,2016,369(2):136-140.
[4]? DEL RIO M,PRETZSCH H,ALBERDI I,et al.Characterization of the structure,dynamics,and productivity of mixed-species stands:review and perspectives[J].European Journal of Forest Research,2016,135:23-49.
[5]? NAKAJIMA T,TATSUHARA S.Silviculture guidelines for developing mixed forests in old coniferous plantations under long-rotation stand density control[J].Forest Science and Technology,2012,8:127-134.
[6]? 王瑞輝.我國人工造林種存在的生態問題及對策[J].湖南林業科技,1999,26(4):30.
[7]? 陳霞,袁在翔,金雪梅,等.紫金山針闊混交林群落特征[J].中國城市林業,2019,17(5):12-17.
[8]? 田大倫,項文化,閆文德.馬尾松與濕地松人工林生物量動態及養分循環特征[J].生態學報,2004,24(10):2207-2210.
[9]? FORRESTER D I.Linking forest growth with stand structure:Tree size inequality,tree growth or resource partitioning and the asymmetry of competition[J].Forest Ecology and Management,2019,447:139-157.
[10] HACKER S D,GAINES S D.Some implications of direct positive interactions for community species diversity[J].Ecology,1997,78(7):1990-2003.
[11] 譚凌照,范春雨,范秀華.吉林蛟河闊葉紅松林木本植物物種多樣性及群落結構與生產力的關系[J].植物生態學報,2017,41(11):1149-1156.
[12] 魯君悅,吳兆飛,張春雨,等.吉林蛟河針闊混交林林層結構對生產力影響研究[J].生態學報,2021,41(5):2024-2032.
[13] 張結存,徐麗華,張茂霞,等.基于物種空間結構和多樣性的改進型混角度研究[J].浙江農林大學學報,2014,31(3):336-342.
[14] 周紅敏,彭輝,蔣敏,等.鳳陽山自然保護區針闊混交林空間結構研究[J].福建林業科技,2019,46(3):51-54,67.
[15] 朱軍,湛斌,陳霞,等.紫金山針闊混交林空間結構特征與植物多樣性分析[J].中南林業科技大學學報,2018,38(1):75-80,93.
[16] 康希睿,李曉剛,張涵丹,等.不同混交措施下杉木人工林群落穩定性特征[J].生態學雜志,2020,39(9):2912-2920.
[17] 鄧紅兵,陳喆菲,許東,等.森林多目標經營單元的時空尺度擴展[J].生態學報,2015,35(1):65-70.
[18] 李建,李曉宇,曹靜,等.長白山次生針闊混交林群落結構特征及群落動態[J].生態學報,2020,40(4):1195-1206.
[19] 王燕,畢潤成,許強.山西太岳山南部針闊混交林群落特征及空間分布格局[J].廣西植物,2017,37(7):901-911.
[20] 高杰,張君,程艷霞,等.長白山不同林型空間格局及樹種多樣性多尺度分析[J].北京林業大學學報,2014,36(6):103-104.
[21] 杜志,亢新剛,包昱君,等.長白山云冷杉林不同演替階段的樹種空間分布格局及其關聯性[J].北京林業大學學報,2012,34(2):14-19.
[22] 石蒙蒙,楊華,王全軍,等.長白山云冷杉針闊混交林幼苗幼樹空間分布與關聯性[J].北京林業大學學報,2020,42(4):1-11.
[23] 李艷麗,楊華,亢新剛,等.長白山云冷杉針闊混交林天然更新空間分布格局及其異質性[J].應用生態學報,2014,25(2):311-317.
[24] 常新華,趙秀海,曾凡勇,等.長白山針闊混交林主要樹種空間分布及其環境解釋[J].北京林業大學學報,2009,31(1):7-12.
[25] 鄧恢.馬尾松闊葉樹混交林土壤理化性質比較研究[J].福建林業科技,2012,39(1):41-44,52.
[26] PANG Y,TIAN J,ZHAO X,et al.The linkages of plant,litter and soil C:N:P stoichiometry and nutrient stock in different secondary mixed forest types in the Qinling Mountains,China[J].Peerj,2020,8:e9274.
[27] 于飛.臨朐縣石灰巖山區針闊混交林土壤理化性狀及水文效應研究[D].泰安:山東農業大學,2014.
[28] 林文樹,穆丹,王麗平,等.針闊混交林不同演替階段表層土壤理化性質與優勢林木生長的相關性[J].林業科學,2016,52(5):17-25.
[29] 秦倩倩,王海燕,李翔,等.長白山云冷杉針闊混交林半分解層凋落物生態功能[J].林業科學研究,2019,32(1):147-152.
[30] BRADFORD M A,BERG B,MAYNARD D S,et al.Understanding the dominant controls on litter decomposition[J].Journal of Ecology,2016,104:229-238.
[31] 朱金兆,劉建軍,朱清科,等森林凋落物層水文生態功能研究[J].北京林業大學學報,2002,24(5):30-34.
[32] 蔣云峰.長白山針闊混交林主要凋落物分解及土壤動物的作用[D].長春:東北師范大學,2013.
[33] 劉世榮.中國森林生態系統水文生態功能規律[M].北京:中國林業出版社,1996.
[34] 常雅軍,曹靖,李建建,等.秦嶺西部山地針葉林凋落物層的化學性質[J].生態學雜志,2009,28(7):1308-1315.
[35] 李小雙,趙安娜,黨承林,等.昆明西山云南油杉針闊混交林的群落結構及其更新特征研究[J].云南大學學報(自然科學版),2013,35(4):549-557.
[36] YAN Y,ZHANG C,WANG Y,et al.Drivers of seeding survival in a temperate forest and their relative importance at three stages of succession[J].Ecology & Evolution,2015,5(19):4287-4299.
[37] SEIWA K,ETOH Y,HISITA M,et al.Roles of thinning intensity in hardwood recruitmentand diversity in a conifer,Criptomeria japonica plantation:a 5-year demographicstudy[J].Forest Ecology & Management,2012,269(2):177-187.
[38] 米爽,宋子龍,秦江環,等.撫育采伐對吉林蛟河針闊混交林幼苗更新的影響[J].北京林業大學學報,2019,41(5):159-169.
[39] GILADI I,ZIV Y,MAY F,et al.Scale-dependent determinants of plant species richness in a semi-arid fragmented agro-ecosystem[J].Journal of Vegetation Science,2011,6:983-996.
[40] HE F L,LAFRANKIE J V,SONG B.Scale dependence of tree abundance and richness in a tropical rain forest,Malaysia[J].Landscape Ecology,2002,17:559-568.
[41] 汪殿蓓,暨淑儀,陳飛鵬,等植物群落物種多樣性研究綜述[J].生態學雜志,2001,20(4):55-60.
[42] 馬志遠,石玲,吳相菊,等.長白山次生針闊混交林喬木多樣性維持機制研究[J].北京林業大學學報,2014,36(6):93-98.
[43] 辛營營,韋新良.青山湖針闊混交林優勢樹種競爭的數量研究[J].浙江農林大學學報,2011,28(4):601-606.
[44] 杜秀芳,湯孟平,潘建勇,等.臨安區不同森林類型競爭指數比較研究[J].生態學報,2020,40(12):4064-4072.
[45] 田超,楊新兵,劉陽.邊緣效應及其對森林生態系統影響的研究進展[J].應用生態學報,2011,22(8):2184-2192.
[46] 奚為民,鐘章成,畢潤成.四川縉云山森林群落林窗邊緣效應的研究[J].植物生態學與地植物學學報,1993(3):42-52.
[47] 劉彥春,張遠東,劉世榮,等.川西亞高山針闊混交林喬木層生物量、生產力隨海拔梯度的變化[J].生態學報,2010,30(21):5810-5820.
[48] 丘菊秀.針闊混交林的優勢及應用分析[J].綠色科技,2020(7):119-121.
[49] 宋德利.人工針闊混交林水源涵養能力的試驗[J].防護林科技,2014(2):36-37,104.
[50] 張光燦,劉霞.林冠截留降雨模型研究進展及其述評[J].南京林業大學學報,2000,24(1):64-68.
[51] 郝新朝,安裕倫,伍顯,等.貴州省森林生態系統水源涵養功能價值評估[J].安徽農學通報,2019,25(Z1):94-97,130.
[52] 劉涌.混交林的優點及混交原則相關問題分析[J].科技創新與應用,2014,(12).
[53] 秦娟,唐紅心,楊雪梅.馬尾松不同林型對土壤理化性質的影響[J].生態環境學報,2013,2(4):598-604.
[54] 陳杰.白樺+紅松人工混交林生態效益的研究[J].防護林科技,2019(9):27-28,36.
[55] 張發根,張文斌,賴根偉,等.混交林下多花黃精仿野生種植技術探討[J].華東森林經理,2019,33(4):22-24.
[56] 臧麗鵬,張廣奇,何躍軍,等.近自然經營對黔中地區馬尾松灌草群落多樣性及物種共存的影響[J].山地農業生物學報.2021,40(3):21-28.
[57] 鐘梁,高友英,孫浩,等.扶河上游生態公益林4種森林類型枯落物層和土壤層水文效應[J].南方林業科學,2017,45(6):5-8.
[58] LIU C L C,KUCHMA O,KRUTOVSKY K.Mixed-species versus monocultures in plantation forestry:Development,benefits,ecosystem services and perspectives for the future[J].Global Ecology and Conservation,2018,15:e00419.
[59] 劉瓊.森林康養產業發展的必然性可行性和路徑選擇[J].經濟師,2021(2):36-37,39.
[60] BRAVO-OVIEDO N,PRETZSCH H,AMMER C,et al.European mixed forests:definition and research perspectives[J].Forest Systems,2014,23(3):518-533.
[61] 于瀚,歐靜,漆倩,張智葉.黔中地區馬尾松-杜鵑植物群落多樣性研究[J].山地農業生物學報,2020,39(2):14-20.
[62] 陳福軍.發展林下經濟推進林下資源可持續發展利用的探索[J].農業開發與裝備,2020(7):39.
[63] 黃美水.談林下經濟產業現狀及發展重點[J].百科論壇電子雜志,2019(16):227.
通訊作者:張健(1980—)男,博士,副教授,主要從事微生物生態學方面研究,E-mail:zhangjian12102@163.com.
猜你喜歡
綠色科技(2016年21期)2016-12-27 16:05:03
現代管理科學(2017年1期)2016-12-26 17:16:55
考試周刊(2016年21期)2016-12-16 10:37:45
當代經濟管理(2016年11期)2016-12-14 17:47:10
出版廣角(2016年14期)2016-12-13 01:49:53
新課程·中旬(2016年9期)2016-12-01 12:56:28
商情(2016年40期)2016-11-28 10:54:26
青年文學家(2016年30期)2016-11-22 19:02:32
科教導刊(2016年27期)2016-11-15 22:36:28
科技視界(2016年18期)2016-11-03 22:02:50
